表1 霍格兰溶液配方
Tab.1 Hoagland solution formulation
钾在植物中含量很高,是植物必需的大量元素之一,在植物的光合作用、气孔运动、蛋白合成和氧化代谢等生命活动中起重要作用[1-3]。缺钾时,会导致植物生长受限和代谢紊乱,严重缺钾时,植物甚至无法正常生长[4]。
研究表明,不同植物的钾含量不同。胡笃敬等[5]研究了30种植物的含钾量,包括水生植物、陆生野生植物、绿肥植物、早稻和晚稻等,结果发现,不同植物间的钾含量差别很大。此外,同一植物的不同品种钾含量也不同,如大豆[6]、燕麦[7]、小麦[8]、香蕉[9]、甘薯[10]、甘蔗[11]、玉米[12]、西瓜[13]、油菜[14]等。烟草品种间的烟叶含钾量,也存在明显差异[15-17]。杨欢等[18]研究了93份烟草种质资源材料的烟叶钾含量,并根据钾含量的高低,将这些种质材料分为高钾基因型、普通型与低钾型。
在分子层面,植物对外界钾的吸收,主要通过高亲和与低亲和2类吸收机制进行[3]。其中,低亲和机制主要是由位于细胞质膜上的钾离子通道承担,是植物吸收钾离子的重要途径之一[19-20]。例如,拟南芥钾离子通道的AKT1,是拟南芥根从外界获取K+的主要组分之一[21]。CIPK是一类植物特有的丝氨酸/苏氨酸蛋白激酶[22],CBL-CIPK信号系统广泛参与了植物钾吸收调节过程[23]。研究表明,在AtCBL1/9的介导下,AtCIPK23通过磷酸化AKT1,激活该离子通道,从而增强了拟南芥在低K+条件下对K+的吸收[24]。在水稻中,OsCBL1-OsCIPK23同样通过调控OsAKT1,参与了水稻的K+吸收[25]。在胡杨中,PeCBL1-PeCIPK23通过磷酸化钾离子通道PeKC1、PeKC2,从而实现其对低钾胁迫的响应[26]。还有研究表明,AtCIPK23在烟草中的过表达导致了转基因烟草低K+耐受性的增加[27]。此外,甘蔗CIPK23[28]、水稻CIPK10[22]、林烟草CIPK3[29]、普通烟草CIPK2等[30]均可能参与了植物对K+的吸收调节过程。
烟草是我国重要的经济作物,烟叶含钾量高低,是衡量烟叶品质的重要指标之一[31]。然而,目前对于烟草钾吸收的分子机制研究尚不够深入,尤其是NtCIPK基因参与钾吸收调控的研究还较少,其调控机制尚不清楚。因此,本研究对3个烤烟品种进行了不同K+浓度的处理,研究了其干质量、钾含量及NtCIPK家族11个基因的表达模式,并分析了这3个品种钾含量和其NtCIPK基因表达之间的关系,旨在为烟草钾吸收调控的分子机制研究提供一定的参考。
参试烤烟品种为K326、贵烟5号、云87。
试验所需的TRIzol试剂、cDNA合成试剂盒、SYBR Green Master mix等试剂购自宝生物工程(大连)有限公司。
1.2.1 不同钾浓度培养溶液的配制 不同钾浓度的培养液,在霍格兰溶液配方基础上进行适当调整配制而成(表1)。用氯化钾调整培养溶液的钾浓度分别为6.0,1.0,0.2 mmol/L,其他不变。
表1 霍格兰溶液配方
Tab.1 Hoagland solution formulation
1.2.2 烟草幼苗的培养与处理 挑选大小一致且饱满的烟草种子,表面消毒后,分别播种在白色泡沫漂浮育苗盘(长65.5 cm、宽33.5 cm、高4.5 cm、162穴)中,然后,将育苗盘放置在育苗池中进行育苗。育苗池中加注K+浓度为6 mmol/L的霍格兰溶液。60 d后,每品种挑选长势一致的幼苗各90株,分为3组,分别放入K+浓度为6.0,1.0,0.2 mmol/L的霍格兰溶液中,进行处理。每个处理包括30株烟苗,10株烟苗为一个重复,共重复3次。
处理48 h后,对每个浓度每个品种的烟苗进行随机取样。取样时,取每个重复烟苗叶片混合样,在液氮中速冻后,置于-80 ℃冰箱中保存,用于RNA的提取。
处理28 d后,将参试的全部烟株取出,并将根部冲洗干净,用吸水纸吸干水分后,在烘箱中105 ℃杀青30 min,80 ℃下烘干至恒质量待用。
1.2.3 烟株干质量及钾含量的测定 用电子天平称取烟株的干质量。植株钾含量的测定采用火焰光度计法[32]进行。
1.2.4 NtCIPK基因的表达模式分析 RNA的提取:采用TRIzol法进行,cDNA的合成参照试剂盒说明,具体参照卓维等[30]的方法进行。
表2 引物序列
Tab.2 Primer sequences

引物设计:根据四川农业大学农学院的分子生物技术验室前期所克隆到的11个NtCIPK基因的cDNA序列,采用Primer Premier 5.0设计qPCR引物(表2)。
qPCR扩增:以反转录后的cDNA为模板,以烟草18S rRNA为内参进行qPCR扩增。qRT-PCR扩增体系为:2×Prime STAR Max 5.0 μL,上游引物 0.5 μL,下游引物 0.5 μL,cDNA 1 μL,ddH2O 3 μL。qRT-PCR扩增程序为:50 ℃反应2 min,95 ℃预变性3 min;然后,95 ℃变性10 s,65 ℃延伸45 s,进行39个循环。基因相对表达水平用2-ΔΔCt方法进行计算。
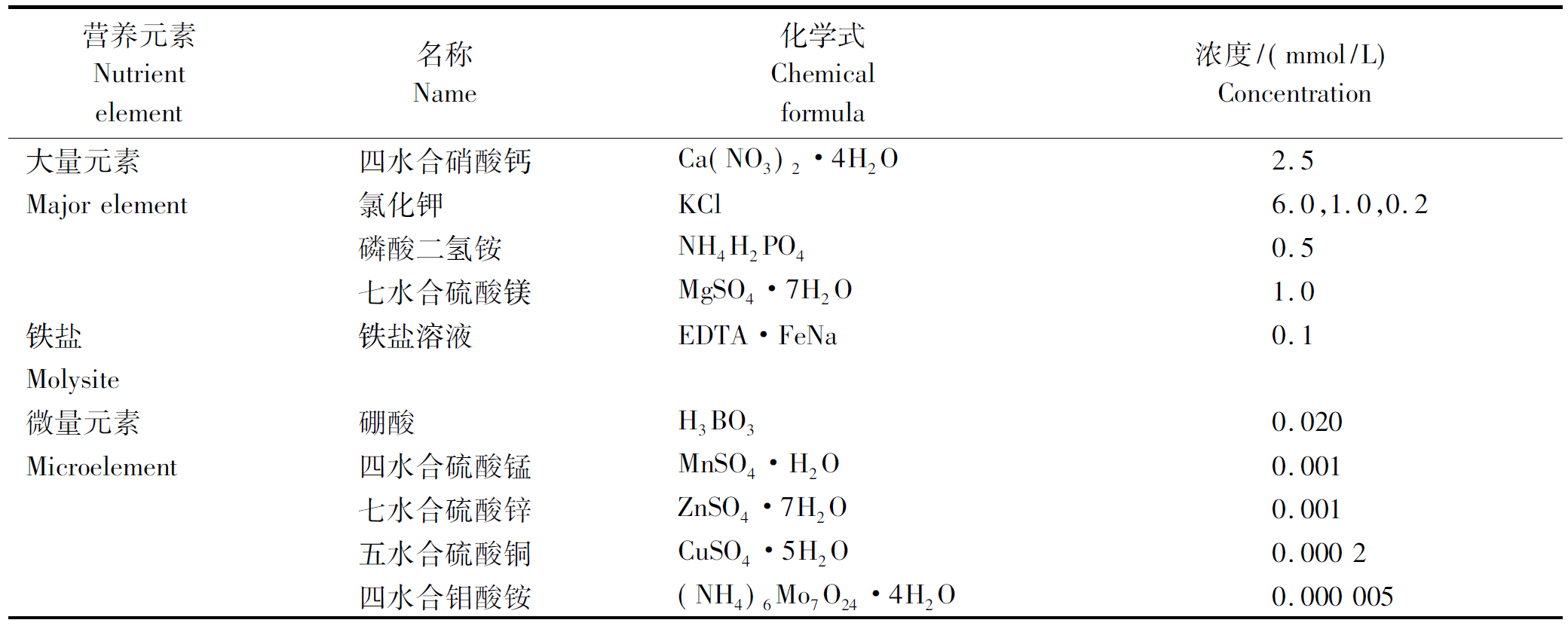
在不同K+浓度处理下,各品种生物量的测定结果如图1所示。随着培养液K+浓度的降低,各品种的植株干质量均呈下降趋势。在1.0,0.2 mmol/L K+浓度处理下,云87与贵烟5号的干质量差异不显著,但在0.2 mmol/L K+浓度处理下云87与贵烟5号的干质量均显著高于K326;在6.0 mmol/L K+浓度处理下,贵烟5号的植株干质量显著低于K326和云87,后二者之间的差异则不显著。烟草植株生物量的测定结果表明,外界供K+量的高低,显著影响到烟草植株的生长与干物质的积累。
图中每个浓度不同品种的不同小写
字母表示在P<0.05水平上差异显著。图2-3同。
Different lowercase letters in each concentration of the figure
indicate significant difference atP<0.05.The same as Fig.2-3.
图1 各品种烤烟不同K+浓度处理的干质量
Fig.1 Dry weight of different varieties of flue-cured tobacco treated with different K+ concentrations
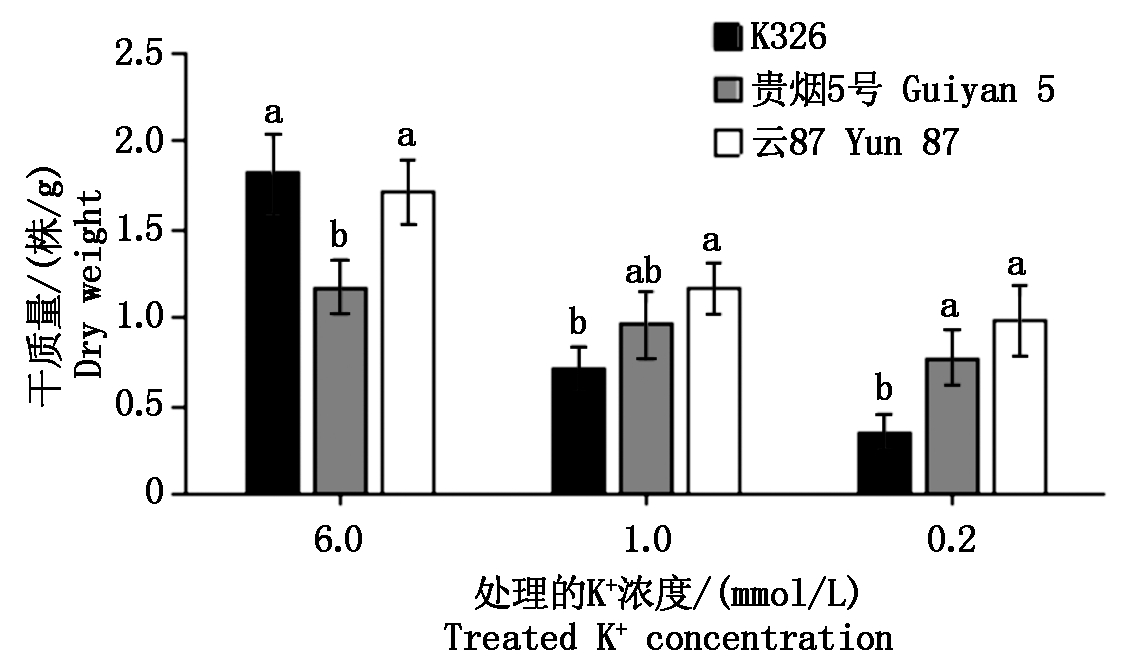
图2为不同K+浓度处理下,各处理烟草植株钾积累量(株/g)测定结果。随培养液中K+浓度的降低,各参试材料植株的钾积累量都呈下降趋势。在6.0,1.0 mmol/L K+浓度处理下,云87的钾积累量显著高于K326及贵烟5号;而在0.2 mmol/L K+浓度处理下,云87与贵烟5号和K326的钾积累量差异显著。钾积累量的测定结果表明,不同烟草品种在不同的外界K+浓度环境中,其植株的K+积累特性明显不同。
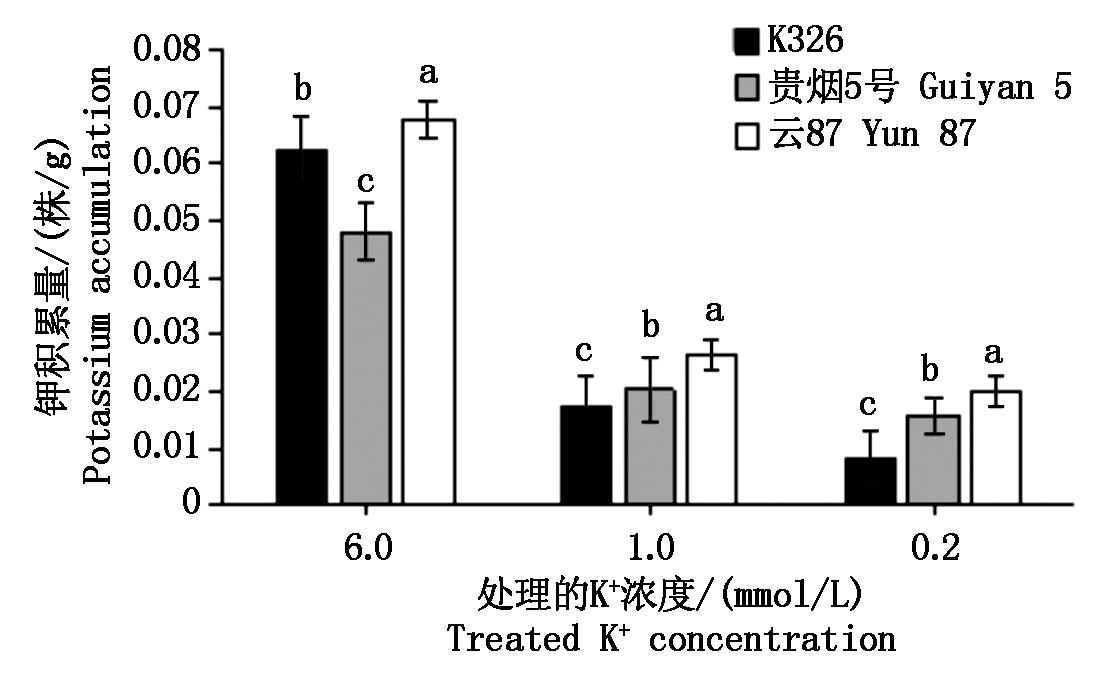
图2 各品种烤烟不同K+浓度处理的K+积累量
Fig.2 K+ accumulation of different K+ concentrations of flue-cured tobacco of different varieties
对在不同K+浓度处理下,11个NtCIPK基因在各烟草品种中的表达进行了测定,结果如图3所示。
从图3可以看出,NtCIPK基因的表达均随着外界钾浓度的变化而改变。在低钾条件下(0.2 mmol/L),其表达量基本是上调的,其中,云87的11个NtCIPK基因表达量均高于对照K326,且存在显著性差异,显示这些基因的表达受低钾胁迫的诱导。但NtCIPK9和NtCIPK24分别在K326及贵烟5号中的表达,随着外界钾浓度的降低,却呈下调趋势,即随着K+浓度的降低而下调,都分别显著低于云87的表达量。在所检测的11个NtCIPK基因中,NtCIPK2、NtCIPK5、NtCIPK6、NtCIPK9、NtCIPK12、NtCIPK14、NtCIPK23和NtCIPK24随着K+浓度的降低,表达量变化较大。可以推测,这8个NtCIPK基因可能以某种直接或间接的方式参与了烟草钾吸收的调控。
图3 不同烤烟品种在不同K+浓度处理后的NtCIPK基因的相对表达量
Fig.3 Relative expression of NtCIPK gene after treatment with different K+ concentrations in different flue-cured tobacco varieties
2.4.1 逐步回归分析 将各处理烟草植株的K+积累量(Y)与11个NtCIPK基因的表达量进行逐步回归分析,得出了3个烟草品种在3个钾浓度处理下的逐步回归方程(表3)。
对K326来说,NtCIPK12(X7)、NtCIPK18(X9)和NtCIPK3(X2)的表达,分别在6.0,1.0,0.2 mmol/L条件下,对其K+积累量影响显著。贵烟5号的情况有所不同,其K+积累量在6.0,1.0 mmol/L条件下与NtCIPK5(X3)的表达密切相关;而在0.2 mmol/L条件下,则受NtCIPK3(X2)表达的显著影响。NtCIPK23(X10)的表达,在外界高K+浓度(6.0 mmol/L)下,与云87植株的K+积累量相关性显著;而在中(1.0 mmol/L)、低钾浓度(0.2 mmol/L)下,云87植株的K+积累量则与NtCIPK9(X5)的表达相关性显著。
2.4.2 通径分析 在回归分析的基础上,对NtCIPK3(X2)、NtCIPK5(X3)、NtCIPK9(X5)、NtCIPK12(X7)、NtCIPK18(X9)及NtCIPK23(X10)做了进一步的通径分析(表4)。
从逐步回归方程和通径分析结果来看,在外界0.2 mmol/L K+浓度下,NtCIPK3(X2)的表达与K326和贵烟5号的植株钾积累量分别呈显著正相关和显著负相关。同时,NtCIPK5(X3)在6.0 mmol/L浓度下的表达,与贵烟5号的植株钾积累量呈显著正相关。此外,在6.0 mmol/L K+浓度处理下,云87的植株钾积累量与NtCIPK23(X10)的表达显著负相关。
表3 各烤烟品种的K+含量和NtCIPK基因表达模式的逐步回归方程
Tab.3 Stepwise regression equations of K+ content and NtCIPK gene expression pattern of various flue-cured tobacco varieties
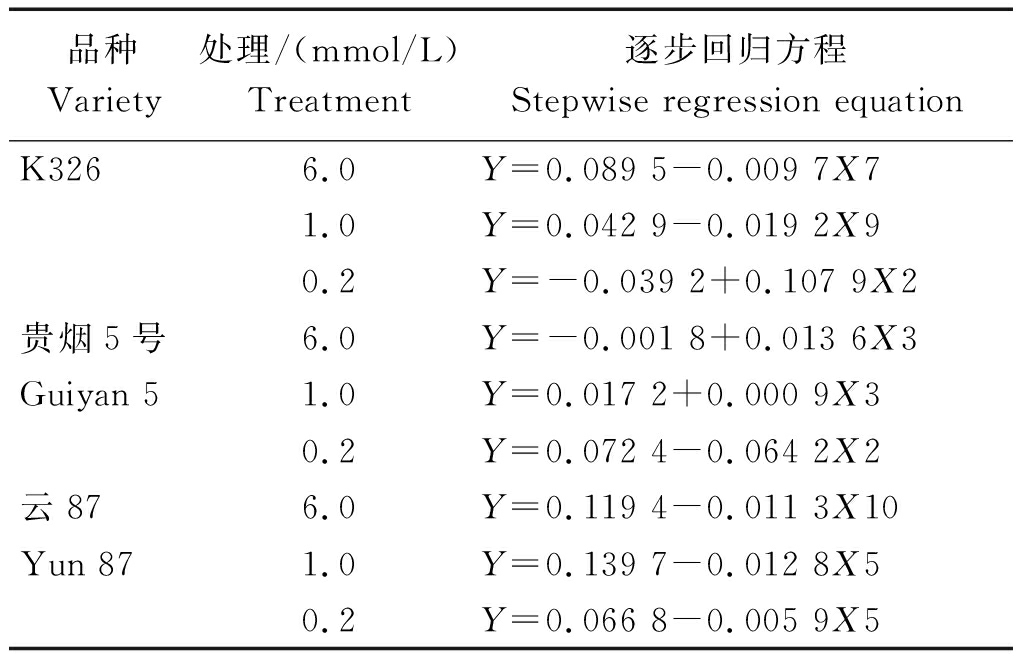
品种Variety处理/(mmol/L)Treatment逐步回归方程Stepwise regression equationK3266.0 Y=0.089 5-0.009 7X71.0Y=0.042 9-0.019 2X90.2Y=-0.039 2+0.107 9X2贵烟5号6.0 Y=-0.001 8+0.013 6X3Guiyan 51.0 Y=0.017 2+0.000 9X30.2Y=0.072 4-0.064 2X2云876.0 Y=0.119 4-0.011 3X10Yun 871.0 Y=0.139 7-0.012 8X50.2Y=0.066 8-0.005 9X5
注: Y.对应各浓度烤烟品种的K+含量;X1、X2、X3、X4、X5、X6、X7、X8、X9、X10、X11分别代表NtCIPK2、NtCIPK3、NtCIPK5、NtCIPK6、NtCIPK9、NtCIPK11、NtCIPK12、NtCIPK14、NtCIPK18、NtCIPK23、NtCIPK24基因的相对表达量。表4同。
Note: Y. The K+ content of the corresponding flue-cured tobacco varieties;X1, X2, X3, X4, X5, X6, X7, X8, X9, X10, and X11 represent relative expression levels ofNtCIPK2,NtCIPK3,NtCIPK5,NtCIPK6,NtCIPK9,NtCIPK11,NtCIPK12,NtCIPK14,NtCIPK18,NtCIPK23, andNtCIPK24 genes, respectively. The same as Tab.4.
表4 各烤烟品种的K+含量和NtCIPK基因表达模式的通径分析
Tab.4 Path analysis of K+ content and NtCIPK gene expression patterns of flue-cured tobacco varieties
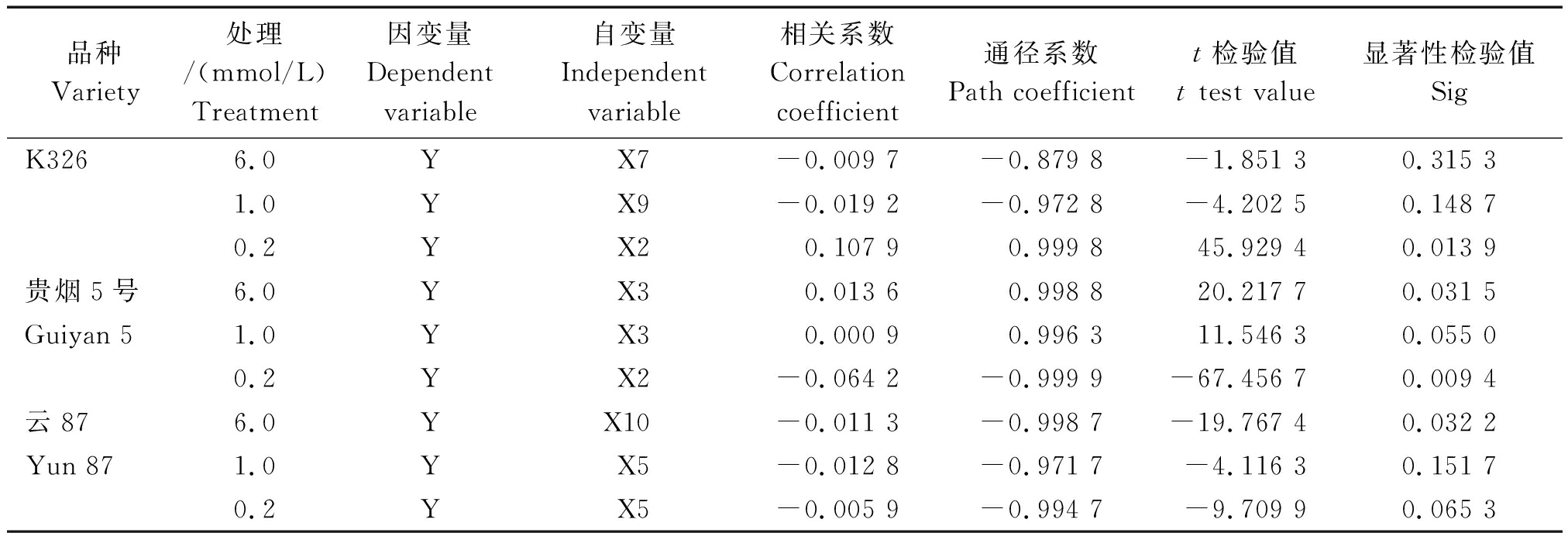
品种Variety处理/(mmol/L)Treatment因变量Dependent variable自变量Independent variable相关系数Correlation coefficient通径系数Path coefficientt检验值t test value显著性检验值SigK3266.0YX7-0.009 7-0.879 8-1.851 30.315 31.0YX9-0.019 2-0.972 8-4.202 50.148 70.2YX20.107 90.999 845.929 40.013 9贵烟5号6.0YX30.013 60.998 820.217 70.031 5Guiyan 51.0YX30.000 90.996 311.546 30.055 00.2YX2-0.064 2-0.999 9-67.456 70.009 4云876.0YX10-0.011 3-0.998 7-19.767 40.032 2Yun 871.0YX5-0.012 8-0.971 7-4.116 30.151 70.2YX5-0.005 9-0.994 7-9.709 90.065 3
本研究通过对3个烟草品种进行不同K+浓度处理,测定了其干质量、钾积累量,并分析了11个NtCIPK基因在各个处理下的表达量,结果表明,NtCIPK基因的表达量与烟草的钾积累量密切相关。
前人研究结果表明,植物的CIPK基因在植物对K+的吸收调控过程中起着重要的作用[23-26]。在本研究中,烟草11个NtCIPK基因在不同品种中的表达量并不相同,并且均能够响应外界钾浓度的变化,这与前人的研究结果相一致[25,28-30]。此结果暗示,烟草的NtCIPK基因广泛参与了烟草对钾的吸收调控过程。
植物的CIPK基因参与钾吸收调控的机制较为复杂。一般情况下,其过表达能够增强植物对钾的吸收[24-27,33 ]。然而,拟南芥AtCIPK9[34]的过表达则导致对低K+敏感;玉米ZmCIPK9[33]的过量表达却提高了钾的吸收能力。由此说明,不同的NtCIPK基因在植物钾吸收过程中的调控机制不尽相同。本研究也发现,从NtCIPK基因的表达模式来看,随着外界钾浓度的降低,其表达可以分为上调及下调2种情况。有9个NtCIPK基因是上调表达,只有K326的NtCIPK9与贵烟5号的NtCIPK24的表达为下调。其中,上调表达的基因,就包括了与前人的研究结果相一致[29-30, 35]的CIPK2、CIPK3、CIPK23基因。烟草NtCIPK基因的这种不同表达模式,在一定程度上说明了其参与烟草钾吸收调控时,存在正调控、负调控或者其他的复杂方式。
逐步回归和通径分析的结果同样说明,在11个NtCIPK基因中,其表达量与烟草钾积累量的相关性存在显著与不显著之别;即使对同一个基因来说,其在不同品种中的作用也不尽相同。例如,同样在0.2 mmol/L K+浓度处理下,NtCIPK3(X2)与K326的K+含量呈显著正相关;然而,与贵烟5号的K+含量则呈显著负相关。该结果同样说明了烟草NtCIPK基因行使钾吸收调控功能的复杂性。
虽然烟草NtCIPK基因广泛参与了烟草对钾的吸收与调控,然而,其参与的方式、调控的模式与功能行使的途径却十分复杂。而每一个NtCIPK基因在其中发挥作用的详细机制,则需更进一步地深入研究。
[1] White P J, Karley A J. Potassium[J]. Cell Biology of Metals and Nutrient, 2010, 17: 199-224.
[2] Lebaudy A, Véry A A, Sentenac H. K+ Channel activity in plants: genes, regulations and functions[J]. FEBS Letters, 2007, 581(12): 2357-2366.
[3] Wang Y, Wu W H. Potassium transport and signaling in higher plants[J]. Annual Review of Plant Biology, 2013, 64(1): 451-476.
[4] Kanai S, Moghaieb R E, El-Shemy H A, et al. Potassium deficiency affects water status and photosynthetic rate of the vegetative sink in green house tomato prior to its effects on source activity[J]. Plant Science, 2011, 180(2): 368-374.
[5] 胡笃敬, 杨敏元, 刘国华. 高钾植物研究[J]. 湖南农业大学学报:自然科学版, 1980(4): 8-16.
[6] 佟晓楠, 佟晓东, 李兴涛, 等. 低钾胁迫对不同耐性大豆品种干物质积累,转移与分配的影响[J]. 江苏农业科学, 2017, 45(12): 56-59.
[7] 王伟伟, 赵攀衡, 齐冰洁, 等. 钾肥对不同燕麦品种产量和品质的影响[J]. 北方农业学报, 2017, 45(2): 42-45.
[8] 邹春琴, 李振声. 钾利用效率不同的小麦品种各生育期钾营养特点[J]. 中国农业科学, 2002, 35(3): 340-344.
[9] 饶宝蓉, 罗海燕, 杜中军, 等. 不同香蕉品种钾离子吸收动力学研究[J]. 热带作物学报, 2009, 30(11): 1612-1617.
[10] 董 月, 安 霞, 张 辉, 等. 不同品种甘薯的生物量累积,养分吸收和分配规律[J]. 江苏农业科学, 2016, 32(2): 313-318.
[11] 郑 超, 李奇伟, 黄振瑞, 等. 不同品种甘蔗对钾素吸收差异性的研究[J]. 热带作物学报, 2011, 32(12): 2221-2225.
[12] 王西志, 王 斌, 李迎春, 等. 不同钾营养效率玉米品种的筛选[J]. 中国土壤与肥料, 2012(1): 64-68, 88.
[13] 吴 萍. 西瓜钾高效品种的筛选及高效吸收机制研究[D]. 南京: 南京农业大学, 2014.
[14] 刘秀秀. 不同钾效率类型油菜干物质积累及钾素吸收利用差异研究[D]. 武汉: 华中农业大学, 2014.
[15] 牛佩兰, 石 屹, 刘好宝, 等. 烟草基因型间钾效率差异研究初报[J]. 烟草科技, 1996, 36(1): 33-35.
[16] 王 勇, 杨 欢, 陈光登, 等. 不同品种烤烟中后期钾素吸收和积累特征差异[J]. 中国烟草科学, 2017, 38(3): 30-36.
[17] 王 勇, 李廷轩, 陈光登, 等. 不同钾基因型烟草吸收和生理生化特性研究[J]. 中国烟草科学, 2017, 38(5): 56-61.
[18] 杨 欢, 王 勇, 李廷轩, 等. 烟草含钾量的基因型差异及钾高效品种筛选[J]. 植物营养与肥料学报, 2017, 23(2): 451-459.
[19] Véry A A, Sentenac H. Molecular mechanisms and regulation of K+ transport in higher plants[J]. Annual Review of Plant Biology, 2003, 54(1): 575-603.
[20] 鲁黎明,杨铁钊. 高等植物K+吸收转运蛋白及其分子调节[J].西北植物学报, 2006, 26(11): 2402-2410.
[21] Han M, Wu W, Wu W H, et al. Potassium transporter KUP7 is involved in K(+) acquisition and translocation inArabidopsis root under K(+)-Limited conditions[J]. Molecular Plant, 2016, 9(3): 437-446.
[22] 章俊丽, 杨 鹃, 张玉满, 等. 水稻OsCIPK10基因的克隆和功能分析[J]. 生物工程学报, 2009, 25(9): 1394-1401.
[23] Luan S, Lan W, Chul Lee S. Potassium nutrition, Sodium toxicity, and calcium signaling: connections through the CBL-CIPK network[J]. Current Opinion in Plant Biology, 2009, 12(3): 339-346.
[24] Xu J, Li H D, Chen L Q, et al. A protein kinase, interacting with two calcineurin B-like proteins, regulates K+ transporter AKT1 inArabidopsis[J]. Cell, 2006, 125(7): 1347-1360.
[25] 李 娟. 水稻钾离子通道OsAKT1及其调控因子参与水稻钾吸收的实验证据[D]. 北京: 中国农业大学, 2014.
[26] 张和臣. 逆境条件下胡杨CBL-CIPK信号途径转导的分子机制研究[D]. 北京: 北京林业大学, 2010.
[27] Xue G, Lu L M, Yang T Z, et al. Enhanced tolerance to Low-K+stress in tobacco plants, that ectopically express the CBL-interacting protein kinaseCIPK23 gene[J]. Czech Journal of Genetics and Plant Breeding, 2016, 52(2): 77-82.
[28] 凌秋平, 曾巧英, 胡 斐, 等. 甘蔗CIPK23基因克隆及对低钾,干旱,盐胁迫的响应[J]. 分子植物育种, 2015,13(6): 1329-1335.
[29] 董连红, 史素娟, 苏玉龙, 等. 林烟草蛋白激酶NsyICIPK3基因的克隆与功能分析[J]. 植物遗传资源学报, 2015, 16(3): 633-639.
[30] 卓 维, 陈 倩, 鲁黎明, 等. 烟草NtCIPK2基因的克隆及表达分析[J]. 浙江农业学报, 2017, 10(8): 1597-1604.
[31] 周冀衡, 汪邓明, 吕国新, 等. 不同烤烟品种对钾素响应能力的研究[J]. 中国烟草科学, 1998, 19(2): 8-12.
[32] 鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000.
[33] 张 翔. 玉米ZmCIPK9及ZmCBL8在非生物胁迫应答中的功能研究[D]. 北京: 中国农业大学, 2016.
[34] Liu L L, Ren H M, Chen L Q, et al. A protein kinase CIPK9 interacts with calcium sensor CBL3 and regulates K+homeostasis under low-K+stress inArabidopsis[J]. Plant Physiology, 2013, 161(1): 226-277.
[35] 闫亚飞. 烟草NtCIPK23基因的克隆,表达分析及转基因过量表达研究[D]. 重庆: 重庆大学, 2012.
鲁黎明(1965-),男,河南正阳人,副教授,博士,硕士生导师,主要从事烟草钾营养及分子生物学研究。
任学良(1976-),男,山西孝义人,研究员,博士,硕士生导师,主要从事烟草遗传育种与分子生物学研究。