水稻是世界上最主要的粮食作物之一,全球有1/2以上的人口以水稻为主食[1]。据统计,我国水稻的种植面积约占世界水稻种植面积的1/5,位居全球第二[2]。土壤盐碱化是制约我国水稻稳定生产的主要环境因素之一。然而,由于气候变暖、工业污染加上错误的灌溉方式等诸多环境及人为因素使土壤盐碱化程度日趋严重,使之逐渐成为阻碍作物高产、威胁粮食安全的主要因素。因此,深入了解水稻在盐碱胁迫下的响应与遗传机制,发掘和利用耐盐碱相关基因,通过育种手段改良品种的耐盐碱性,是提高盐碱地生产力的一项经济有效的措施[3]。黑龙江省是我国水稻的主产区之一,其粳稻品种的种植面积居全国首位[4]。然而,黑龙江省盐碱地面积达134万hm2,占黑龙江省耕地面积的7%[5]。所以开展黑龙江省盐碱地水稻种植对于盐碱地改良及国家粮食安全均有重要意义。黑龙江省地处松嫩平原,盐碱土壤以苏打盐碱土为主,土壤成分复杂,主要成分为碳酸钠和碳酸氢钠[6]。前人大多数直接利用氯化钠和碳酸钠水溶液对水稻进行盐碱胁迫的模拟试验,解析水稻耐盐碱性[7]。相对而言,利用苏打盐碱土进行胁迫研究更具有实际意义。
近些年来,由于水稻直播具有劳动强度低,省工、省本、省秧田、高效等优点[8],在滨海盐渍土及东北苏打盐碱土稻区甚至全球范围内迅猛发展,已经成为一种重要的栽培方式[9]。水稻的各个生长发育阶段都会受到盐碱土壤的危害,而与其他发育时期不同的是,盐碱土稻区的水稻直播田在水稻芽期就已经开始产生不同程度的盐碱危害,通常会降低水稻的成苗率,破坏有效光合群体的建立,进而导致水稻产量的降低。因此,研究水稻芽期耐盐碱性遗传机制,发掘和利用耐盐碱相关基因,培育适合苏打盐碱土直播的水稻新品种对于提高盐碱土生产力具有重要意义。目前,关于水稻种子的耐盐碱性鉴定及QTL定位均是在NaCl、Na2CO3或NaHCO3水溶液胁迫下考察种子的萌发能力[10],即水稻的发芽期耐盐碱性,而在实际水稻生产中往往是将水稻种子催芽后应用于直播生产或育苗移栽。因此,研究苏打盐碱胁迫下水稻幼芽细胞维持生活,进而诱发成绿苗的能力更具有实际意义,而针对东北地区苏打盐碱胁迫下的水稻芽期耐盐碱鉴定及基因挖掘尚无报道。
在21世纪初期,Remington等[11]首次成功地将关联分析引入植物。到目前为止,关联分析已经被广泛应用于玉米、拟南芥、水稻和大豆等众多植物中[12-15],且在植物的耐盐性研究中已有报道[16-17]。随着水稻全基因组测序的完成,关联分析的方法在发掘水稻数量性状基因的广度和精度等方面体现出巨大的优势[6],已成为基因组学研究的热点之一。但到目前为止,利用关联分析的方法进行水稻芽期耐盐碱研究还鲜有报道。
基于以上原因,本研究以176份粳稻组成的自然群体为试验材料,采用黑龙江省大庆地区中度苏打盐碱土进行芽期耐盐碱性鉴定,利用154对SSR标记进行关联分析,以期获得与苏打盐碱胁迫下显著关联的位点及相应载体材料,为培育适合盐碱地种植的水稻品种提供理论依据。
1 材料和方法
1.1 供试材料
以176份具有广泛地理分布的粳稻品种为材料(表1)。其中有15份材料来自中国辽宁,30份材料来自中国吉林,58份材料来自中国黑龙江,45份材料来自日本,7份材料来自朝鲜,7份来自俄罗斯,14份材料来自韩国。
1.2 粳稻芽期耐盐碱性鉴定
本试验在东北农业大学农学院水稻研究所实验室进行。采用大庆地区中度苏打盐碱土作为胁迫处理,土壤基本情况见表2。每份材料选取50粒饱满的种子放入45 ℃恒温箱中,处理48 h,使种子打破休眠,用3%的次氯酸钠溶液浸泡30 min,用自来水冲洗3次后,放入蒸馏水中进行催芽,以种子芽长等于种子长度1/2为发芽标准。将50粒种子播入已经加好20 g盐碱土的培养皿中放入30 ℃的恒温培养箱中,试验重复3次。分别于第13天和第18天每个品种收获地上部分,用蒸馏水浸洗后置于80 ℃烘箱烘干48 h至恒重,分析天平称重后用100 mmol/L醋酸于90 ℃恒温水浴箱中提取2 h,其间晃动一次,用蒸馏水稀释至一定倍数,用M410型火焰光度计测定地上部Na+浓度(Shoot Na+ concentration,SNC)和K+浓度(Shoot K+ concentration,SKC) ,并进一步计算地上部Na+/K+ (Shoot Na+/K+ radio,SNK),同时在处理第18天时,参照水稻盐害评价体系的标准划分每个品种的盐碱害级别[18](表3)。
表1 176份材料名称与来源
Tab.1 Name and source of the 176 accessions
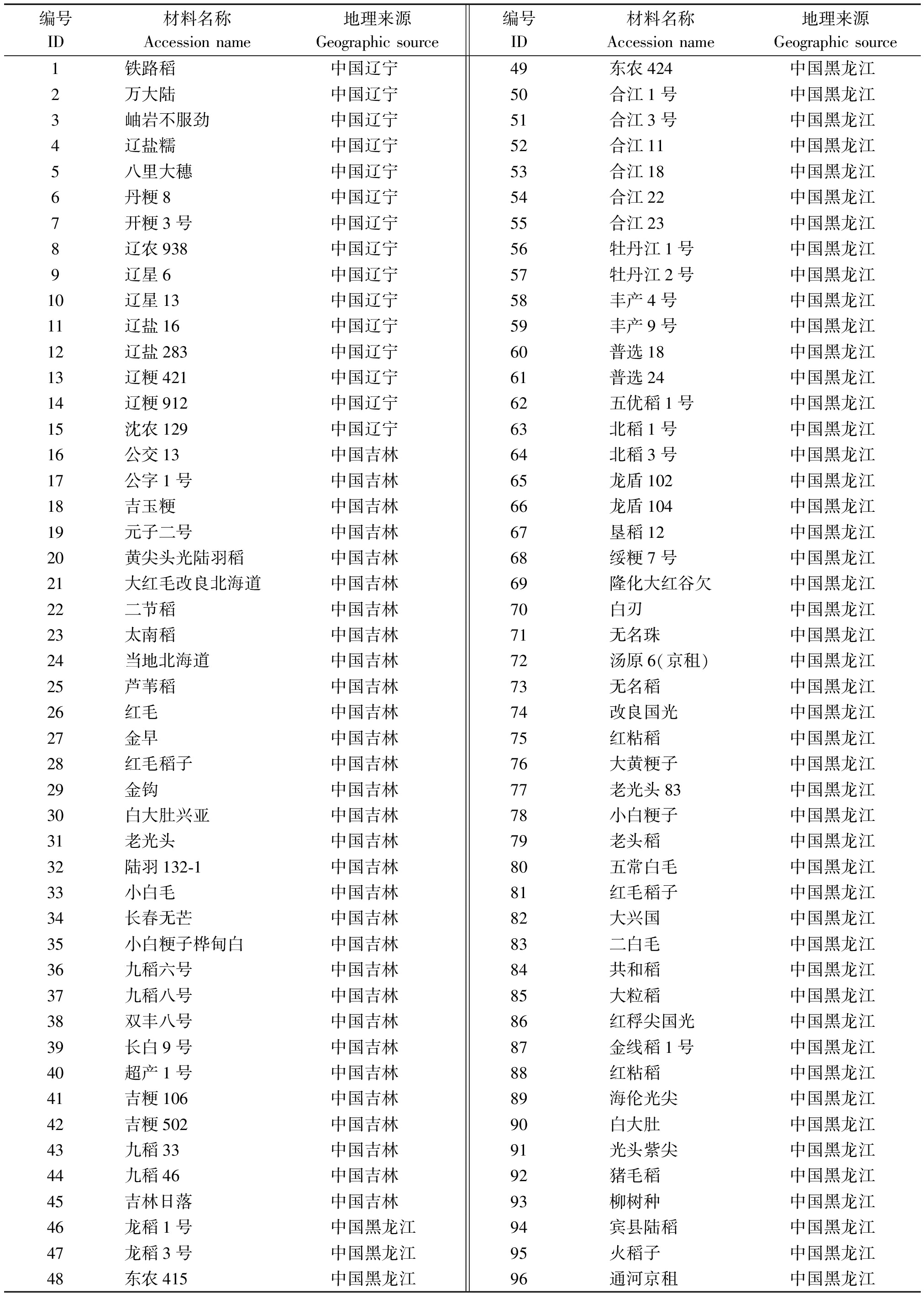
编号ID材料名称Accessionname地理来源Geographicsource编号ID材料名称Accessionname地理来源Geographicsource1铁路稻中国辽宁2万大陆中国辽宁3岫岩不服劲中国辽宁4辽盐糯中国辽宁5八里大穗中国辽宁6丹粳8中国辽宁7开粳3号中国辽宁8辽农938中国辽宁9辽星6中国辽宁10辽星13中国辽宁11辽盐16中国辽宁12辽盐283中国辽宁13辽粳421中国辽宁14辽粳912中国辽宁15沈农129中国辽宁16公交13中国吉林17公字1号中国吉林18吉玉粳中国吉林19元子二号中国吉林20黄尖头光陆羽稻中国吉林21大红毛改良北海道中国吉林22二节稻中国吉林23太南稻中国吉林24当地北海道中国吉林25芦苇稻中国吉林26红毛中国吉林27金早中国吉林28红毛稻子中国吉林29金钩中国吉林30白大肚兴亚中国吉林31老光头中国吉林32陆羽132⁃1中国吉林33小白毛中国吉林34长春无芒中国吉林35小白粳子桦甸白中国吉林36九稻六号中国吉林37九稻八号中国吉林38双丰八号中国吉林39长白9号中国吉林40超产1号中国吉林41吉粳106中国吉林42吉粳502中国吉林43九稻33中国吉林44九稻46中国吉林45吉林日落中国吉林46龙稻1号中国黑龙江47龙稻3号中国黑龙江48东农415中国黑龙江49东农424中国黑龙江50合江1号中国黑龙江51合江3号中国黑龙江52合江11中国黑龙江53合江18中国黑龙江54合江22中国黑龙江55合江23中国黑龙江56牡丹江1号中国黑龙江57牡丹江2号中国黑龙江58丰产4号中国黑龙江59丰产9号中国黑龙江60普选18中国黑龙江61普选24中国黑龙江62五优稻1号中国黑龙江63北稻1号中国黑龙江64北稻3号中国黑龙江65龙盾102中国黑龙江66龙盾104中国黑龙江67垦稻12中国黑龙江68绥粳7号中国黑龙江69隆化大红谷欠中国黑龙江70白刃中国黑龙江71无名珠中国黑龙江72汤原6(京租)中国黑龙江73无名稻中国黑龙江74改良国光中国黑龙江75红粘稻中国黑龙江76大黄粳子中国黑龙江77老光头83中国黑龙江78小白粳子中国黑龙江79老头稻中国黑龙江80五常白毛中国黑龙江81红毛稻子中国黑龙江82大兴国中国黑龙江83二白毛中国黑龙江84共和稻中国黑龙江85大粒稻中国黑龙江86红稃尖国光中国黑龙江87金线稻1号中国黑龙江88红粘稻中国黑龙江89海伦光尖中国黑龙江90白大肚中国黑龙江91光头紫尖中国黑龙江92猪毛稻中国黑龙江93柳树种中国黑龙江94宾县陆稻中国黑龙江95火稻子中国黑龙江96通河京租中国黑龙江
表1(续)

编号ID材料名称Accessionname地理来源Geographicsource编号ID材料名称Accessionname地理来源Geographicsource97有芒紫叶稻中国黑龙江98白芒稻中国黑龙江99新开种中国黑龙江100合江17号中国黑龙江101普通陆稻中国黑龙江102无芒紫叶稻中国黑龙江103丁旭稻中国黑龙江104一目惚日本105关山日本106花脸日本107花水稻日本108绢光日本109千代锦日本110田泰日本111早沙粳日本112分手光日本113富士光日本114山形86日本115WD⁃12468日本116阿利西恩日本117克里斯塔日本118巴尔多日本119珍品稻日本120里歌日本121陆羽日本122青春号日本123藤系168日本124藤系180日本125藤系144日本126岛光日本127功穗日本128秋光日本129野畦日本130屉锦日本131屉时雨日本132秋丰日本133屉稔日本134秋昭日本135清锦日本136藤坂5号日本137陆奥穗秋波日本138越实日本139黄白光日本140西格隆日本141链春号日本142妙妙系原日本143黄金梦日本144早黄金日本145陆奥香日本146小田代5号日本147走坊主日本148道北45日本149平壤1号朝鲜150平壤10号朝鲜151咸南1号朝鲜152咸南23号朝鲜153盐州1号朝鲜154龙城14号朝鲜155朝鲜稻朝鲜156DJP6217俄罗斯1573πP6213(白尖)俄罗斯15811195俄罗斯15912324俄罗斯16012338俄罗斯16112346俄罗斯162X1俄罗斯163云头稻韩国164太星稻韩国165巨楼韩国166敦内韩国167万安韩国168文长韩国169三百韩国170尚美韩国171五台韩国172云长韩国173印月韩国174红珍珠韩国175朝林韩国176早晋富韩国
表2 苏打盐碱土壤基本情况
Tab.2 Soil basic situation of soda saline-alkali

pH全盐含量/‰TotalsaltcontentCO32-/(mg/kg)HCO3-/(mg/kg)SO42-(mg/kg)Na+/(mg/kg)K+/(mg/kg)Ca2+/(mg/kg)8.95.9457382628.21231.6132.310.5
表3 水稻盐碱害评价标准
Tab.3 Evaluation criteria for salt and alkali damage in rice

盐碱害级别Score盐碱害表现Observation耐性Tolerate1植株正常生长,叶片没有受害症状高抗3植株几乎正常生长,但叶尖和少数叶片发白、卷曲抗 5生长严重受阻,大部分植株叶片发生卷曲,只有少数叶片还在伸展中抗7植株完全停止生长,多数叶片干枯,一些植株出现死亡感 9几乎所有植株死亡或接近死亡高感
1.3 DNA提取及SSR标记分析
按照CTAB法提取DNA[19],稍有改进。参照http://www.gramene.org中的引物数据,选择均匀分布于水稻12条染色体上的SSR引物1 000对,由博仕生物技术有限公司合成。利用154个具有多态性的SSR标记进行关联分析。
PCR反应体系为20 μL[20]。扩增结果采用6%聚丙烯酰胺凝胶电泳及银染法检测[21]。
1.4 数据分析
1.4.1 表型数据基本统计分析 利用Excel 2010软件计算性状的平均值、标准差及变异系数等表型数据。
1.4.2 群体结构分析 使用STRUCTURE 2.2软件进行群体结构分析[22]。如果对数似然值 lnP(D) 随亚群数 K 值的增大而增大,则采用 ΔK 来确定合适的 K 值。ΔK 的计算方法是 | L″(K) | 的平均数除以 L′(K) 的标准差,其中,L′(K) = L(K)-L(K-1),| L″(K) | = | L′(K+1) -L′(K) |,STRUCTURE 2.2软件运行结果中的lnP(D)值即为L(K)[22-23]。
1.4.3 连锁不平衡分析 使用D′ 值和r2值作为衡量标记位点间连锁不平衡( Linkage disequilibrium,LD) 程度的标准。其中稀有等位变异(等位变异频率<5%)作为缺失处理。r2值的理论变化范围为0~1,LD越高,连锁越紧密。
1.4.4 耐盐碱相关性状与SSR标记的关联分析 运用TASSEL 3.0软件中的GLM和MLM模型分别进行性状与标记的关联分析。将STRUCTURE 2.2软件运行后形成的Q值作为协变量,然后利用标记变异分别对水稻耐盐碱相关性状的表型值逐一进行回归分析,找出与性状关联的SSR标记位点[22]。当P≤0.01时认为该标记与目标性状关联。
1.4.5 优异等位变异挖掘 在关联分析的基础上,参照Zhang等[24]提出的方法进行优异等位变异的挖掘。以群体平均值为对照,估算关联位点等位变异的表型效应值(表型效应值的计算方法为优异等位基因的性状平均值与群体平均值的差值[25]),同时列出携带该等位基因的5个最佳品种。
2 结果与分析
2.1 苏打盐碱胁迫下粳稻芽期耐盐碱性鉴定
分别对176份粳稻种质的SNC、SKC、SNK及SDG等芽期耐盐碱相关性状进行统计分析(表4)。可以看出,176份粳稻种质资源13 d的SNC、SKC和SNK的3个指标平均值低于18 d的取样结果。18 d的取样结果变异范围和变异系数均大于13 d的取样结果。其中,18 d SKC变异系数最大,达到46.55%;18 d SNC变异范围最大,达到6.78~115.44。从性状的极值来看,品种间的变异范围较大,多样性较为丰富。对数据进行正态分布的适合性检验,发现除SNK以外,其他各个性状的偏度和峰度的绝对值均小于1,说明这些性状基本符合正态分布,呈现典型的数量性状遗传模式。同时,对比表型数据可以得知,随着处理时间增加,钠离子浓度、钾离子浓度及钠钾比均呈现增长趋势。
表4 自然群体耐盐碱性表现
Tab.4 Salt tolerance and alkaline expression in natural population
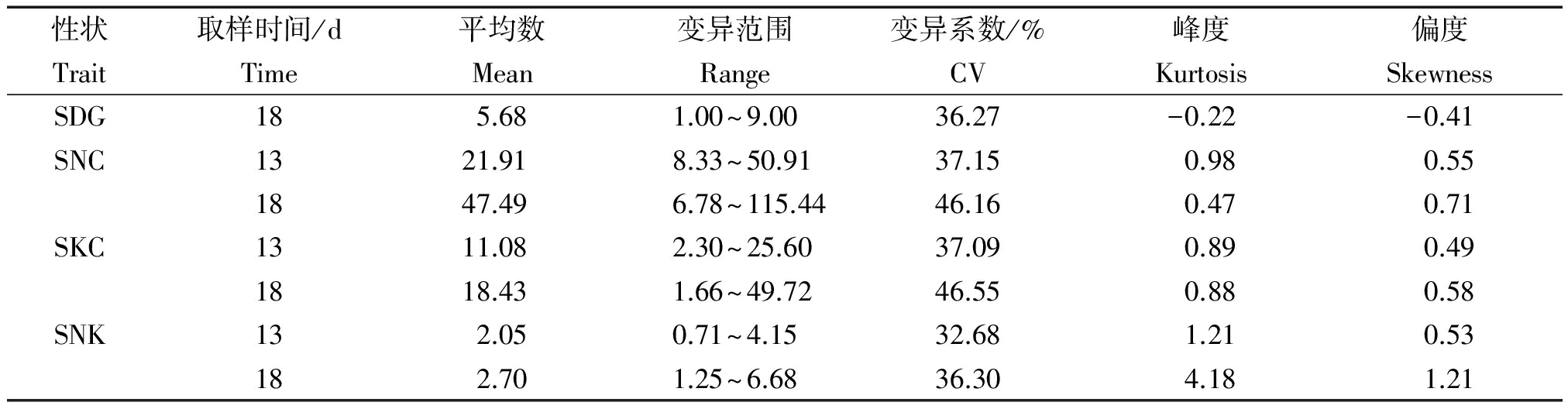
性状取样时间/d平均数变异范围变异系数/%峰度偏度TraitTimeMeanRangeCVKurtosisSkewnessSDG185.681.00~9.0036.27-0.22-0.41SNC1321.918.33~50.9137.150.980.551847.496.78~115.4446.160.470.71SKC1311.082.30~25.6037.090.890.491818.431.66~49.7246.550.880.58SNK132.050.71~4.1532.681.210.53182.701.25~6.6836.304.181.21
2.2 群体结构分析
利用176个水稻品种在154对SSR引物扩增下的基因型数据,结合STRUCTURE 2.2软件对该自然群体进行群体结构分析。结果对数似然值LnP(D)随亚群数K增大而增大,无法确定最适宜的亚群数。因而,采用ΔK方法分析(图1)。当K=3时,ΔK达到最大值,因此,判断样本亚群数为3,分别命名为P1、P2、P3(图2)。采用STRUCTURE软件计算出每个品种归属于3个亚群的后验概率值,以此来划分每个品种所归属的亚群。P1包括47份材料,P2包括71份材料,P3包括14份材料,还有44份材料被划入混合组(MIX)。K为3时各材料的Q值被用于下一步关联分析。
2.3 连锁不平衡分析
使用 TASSEL 3.0 软件对上述3个亚群154个SSR标记位点间的连锁不平衡状况进行了分析(图3),计算出所有可能位点组合的D′值和r2值,用于评价群体的连锁不平衡程度,其中稀有等位变异(等位变异频率<5%)作为缺失处理。评价位点间存在显著的连锁不平衡的标准是P≤0.01。图3用于直观显示处于 LD 状态的位点组合。用不同的颜色表示每对多态性位点之间的D′值及Fisher精确检验的P值(显著性),连锁不平衡现象越明显则颜色更偏向红色,图3中右下角为P值,右上角为D′。发现无论是共线(同一染色体上)SSR位点之间,还是非共线(不同染色体间)SSR标记位点之间均存在一定程度的连锁不平衡。
2.4 粳稻芽期相关耐盐碱性状与SSR标记位点的关联分析
通过TASSEL软件的GLM和MLM模型分别对胁迫13,18 d的耐盐碱相关性状进行关联分析。检测到的与4个耐盐碱相关性状显著关联的SSR位点共有13个,累计34次显著关联(P≤0.01),分布在除第2,3,9,12外的8条染色体上(表5)。共有8个位点在GLM和MLM模型中同时被检测到。
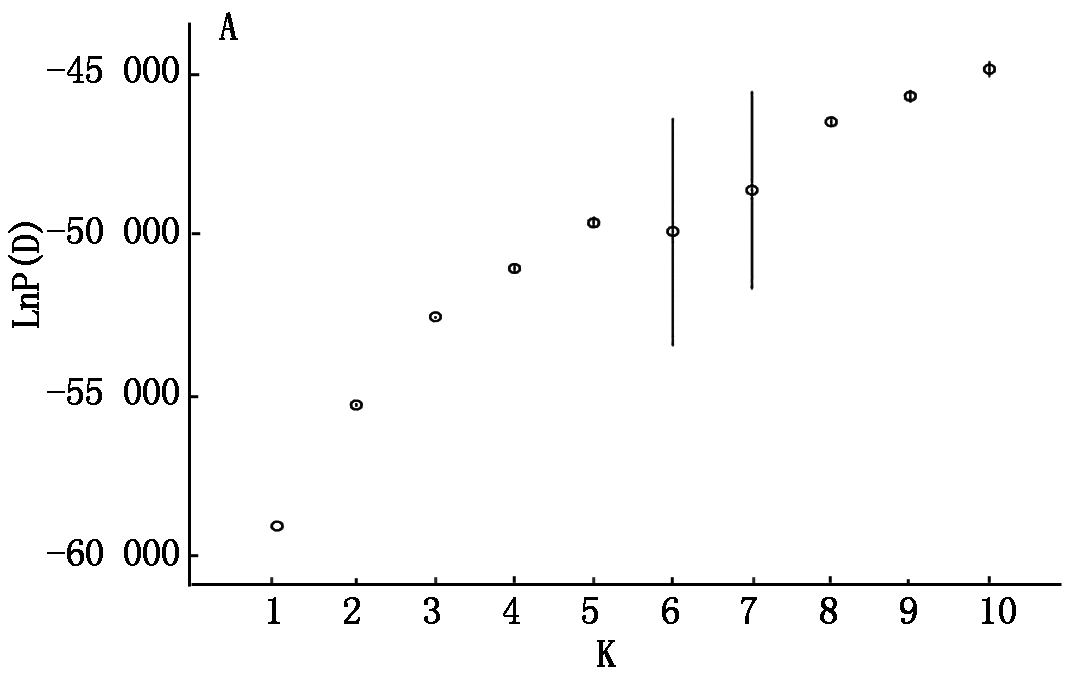
LnP(D)随亚群数K的变化LnP(D)for K from 1 to 10
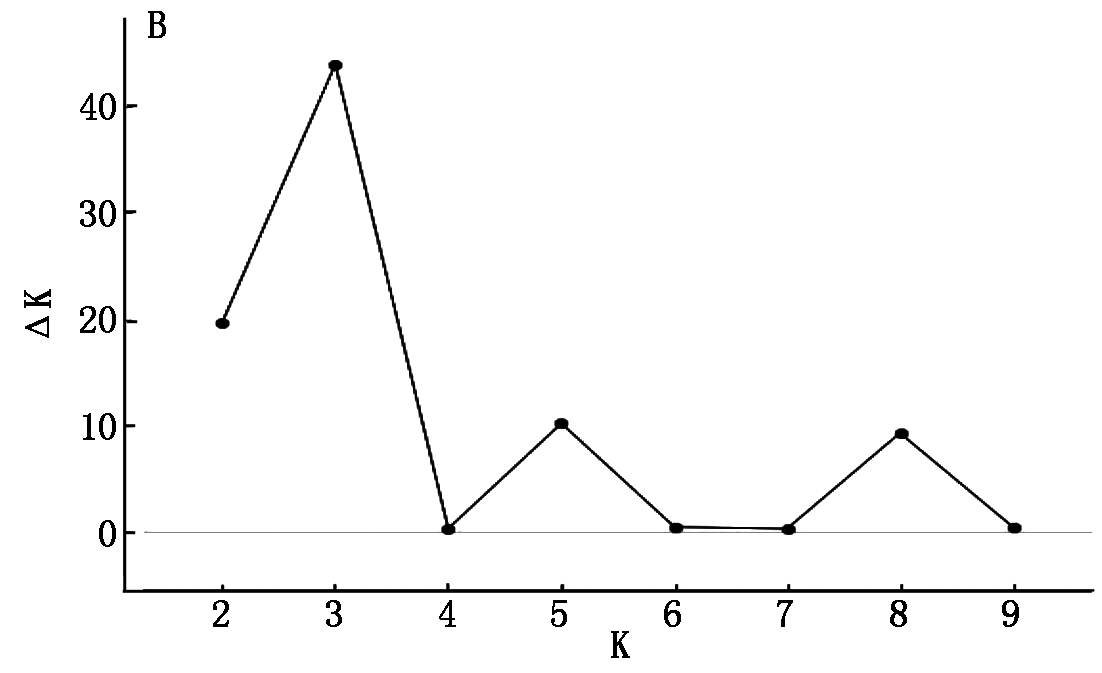
ΔK随亚群数K值的变化曲线ΔK for K from 2 to 10
图1 STRUCTURE软件预测的LnP(D)和ΔK
Fig.1 STRUCTURE software predicts LnP (D) and ΔK
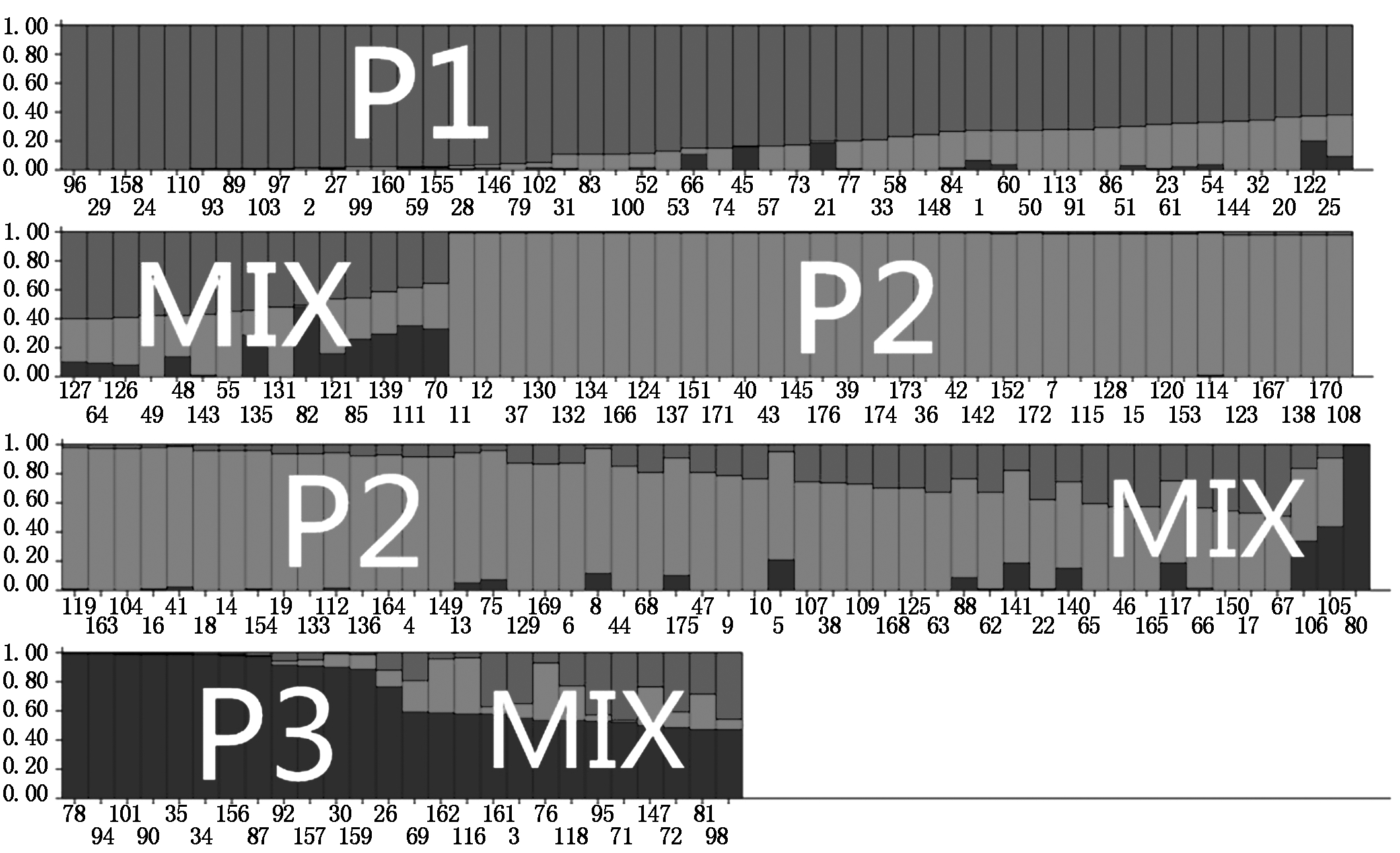
图2 STRUCTURE 2.2检测的群体结构
Fig.2 STRUCTURE 2.2 detection of the population structure

图3 154个SSR位点间的连锁不平衡分布
Fig.3 Linkage disequilibrium between 154 SSR locis
胁迫13 d条件下,共检测到与耐盐碱相关性状显著关联SSR标记6个,在2种模型下均检测出的标记有4个,其中,与SNC-13相关联的标记为RM471,位于第4条染色体上,在GLM和MLM 2种模型的贡献率分别为6.37%,9.30%;在2种模型下同时检测到与SKC-13显著关联的标记有2个,分别为位于第1,6染色体的RM1254和RM527;与SNK-13相关联的标记为位于第11条染色体上的RM1219,在GLM和MLM 2种模型的贡献率分别为6.88%,9.62%。
胁迫18 d条件下,共检测到与耐盐碱相关性状显著关联的SSR标记有10个(累计14次显著关联),在2种模型下同时检测出的标记有7个(累计10次显著关联),其中,与SDG-18显著关联的位点有2个,分别是位于第5和第11染色体上的RM200和RM1219;与SNC-18显著关联的标记有3个,分别是位于第4染色体上的RM241和RM471,及位于第11染色体上的RM286;与SKC-18显著关联的标记有2个,分别是位于第6和第11染色体上的RM527和RM286;与SNK-18相关联的标记有3个,分别是位于第11染色体上的RM286和RM1219,以及位于第10染色体上的RM228。
在2种模型下共检测到4个位点同时与2个或2个以上性状显著关联。分别是与SKC-13和SKC-18关联的RM527;与SNC-13和SNC-18显著关联的RM471;与SNK-13、SNK-18和SDG-18显著关联的RM1219;与SKC-18、SNC-18和SNK-18相关联的RM286。
2.5 优异等位基因及载体材料
将本研究在2种模型中同时检测到的8个位点(累计14次显著关联)进一步挖掘到24个优异等位变异,同时列出携带该等位变异的5个最佳品种(表6)。对于地上部钠离子浓度、地上部钠钾比和盐碱害级别这3个性状来说,优异等位变异的性状平均值要低于群体平均值,所以表型效应值为负数,而对地上部钾离子浓度来说,优异等位变异的性状平均值要高于群体平均值,所以表型效应值为正数。以下载体材料具有耐盐碱的优异等位变异,可作为育种中间材料进行水稻耐盐碱种质资源改良。
表5 苏打盐碱土胁迫下与粳稻芽期耐盐碱性相关性状的相关位点及表型变异的解释率
Tab.5 The explanation of the related loci and phenotypic variation of salt-tolerance-related traits during Japonica rice seedling stage under the stress of soda saline soil
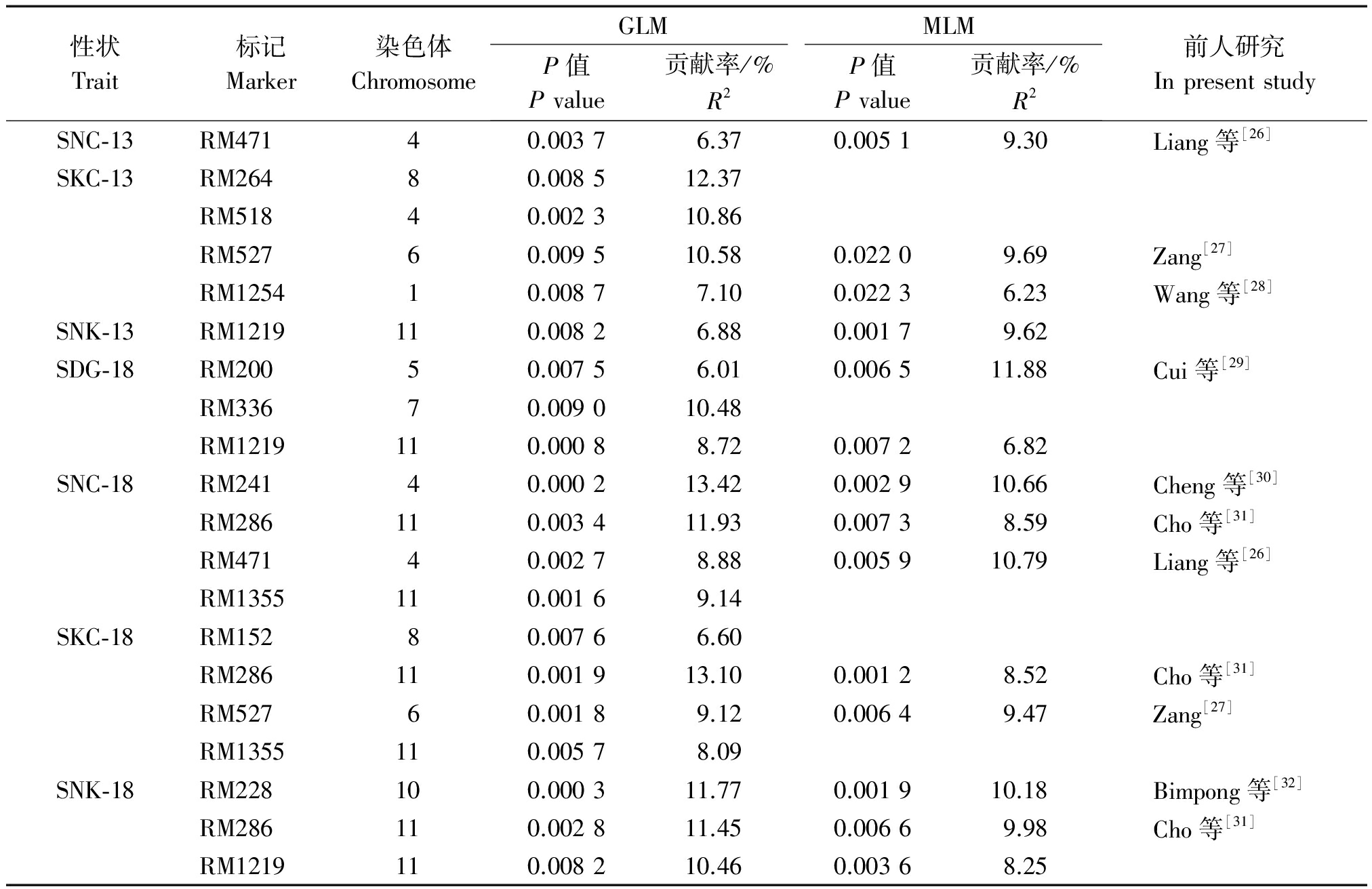
性状Trait标记Marker染色体ChromosomeGLMMLMP值Pvalue贡献率/%R2P值Pvalue贡献率/%R2前人研究InpresentstudySNC⁃13RM47140.00376.370.00519.30Liang等[26]SKC⁃13RM26480.008512.37RM51840.002310.86RM52760.009510.580.02209.69Zang[27]RM125410.00877.100.02236.23Wang等[28]SNK⁃13RM1219110.00826.880.00179.62SDG⁃18RM20050.00756.010.006511.88Cui等[29]RM33670.009010.48RM1219110.00088.720.00726.82SNC⁃18RM24140.000213.420.002910.66Cheng等[30]RM286110.003411.930.00738.59Cho等[31]RM47140.00278.880.005910.79Liang等[26]RM1355110.00169.14SKC⁃18RM15280.00766.60RM286110.001913.100.00128.52Cho等[31]RM52760.00189.120.00649.47Zang[27]RM1355110.00578.09SNK⁃18RM228100.000311.770.001910.18Bimpong等[32]RM286110.002811.450.00669.98Cho等[31]RM1219110.008210.460.00368.25
注:SNC-13、SKC-13、SNK-13代表13 d取样结果;SDG-18、SNC-18、SKC-18、SNK-18代表18 d取样结果。表6同。
Note:SNC-13,SKC-13,SNK-13 represent 13 d sampling results;SDG-18,SNC-18,SKC-18,SNK-18 represent 18 d sampling results.The same as Tab.6.
表6 苏打盐碱土胁迫下与粳稻芽期耐盐碱性相关的优异等位变异及载体材料
Tab.6 Salinity and alkali-tolerant excellent allelic variation and carrier materials during Japonicarice seedling stage under the stress of soda saline soil
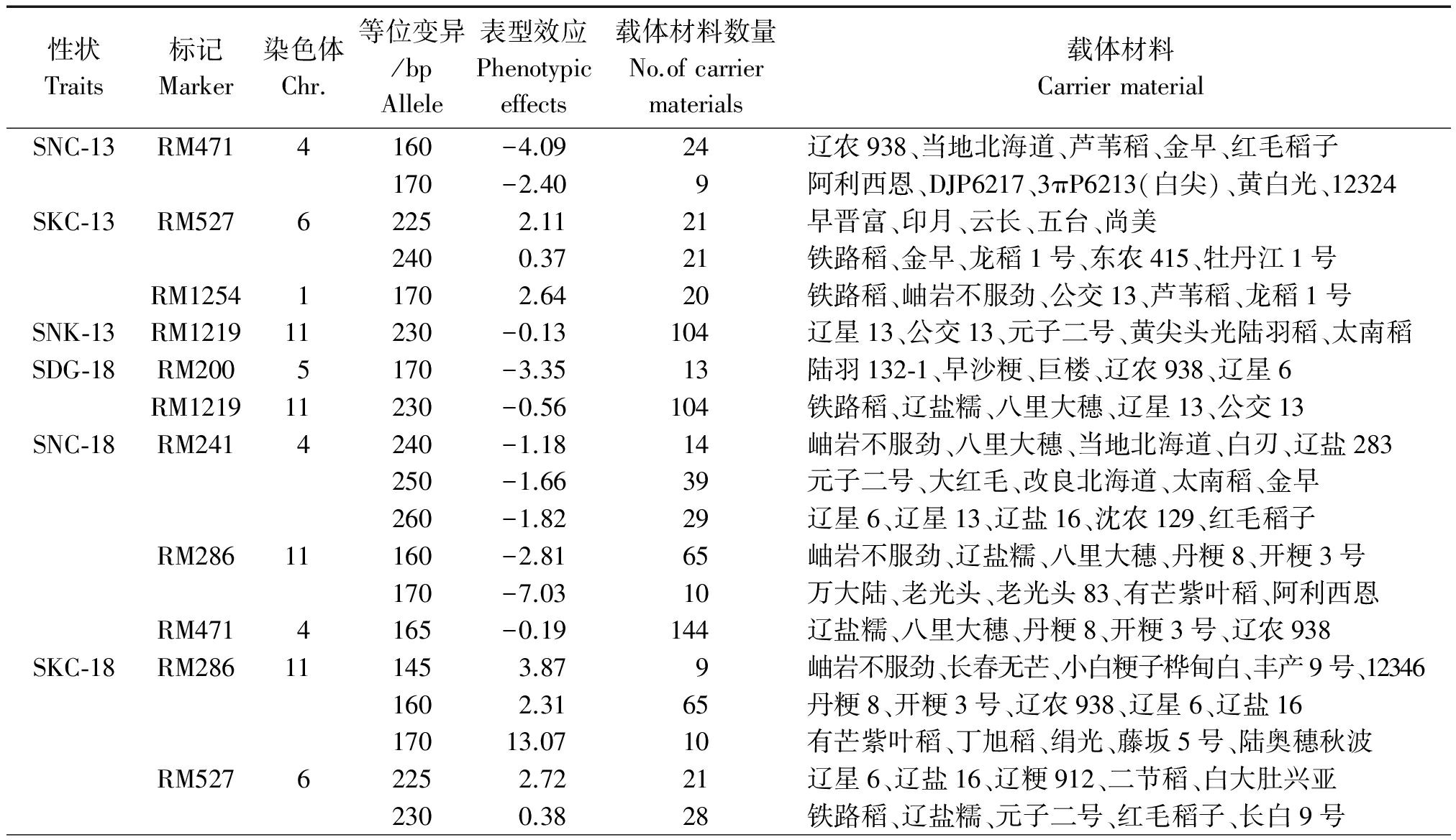
性状Traits标记Marker染色体Chr.等位变异/bpAllele表型效应Phenotypiceffects载体材料数量No.ofcarriermaterials载体材料CarriermaterialSNC⁃13RM4714160-4.0924辽农938、当地北海道、芦苇稻、金早、红毛稻子170-2.409阿利西恩、DJP6217、3πP6213(白尖)、黄白光、12324SKC⁃13RM52762252.1121早晋富、印月、云长、五台、尚美2400.3721铁路稻、金早、龙稻1号、东农415、牡丹江1号RM125411702.6420铁路稻、岫岩不服劲、公交13、芦苇稻、龙稻1号SNK⁃13RM121911230-0.13104辽星13、公交13、元子二号、黄尖头光陆羽稻、太南稻SDG⁃18RM2005170-3.3513陆羽132⁃1、早沙粳、巨楼、辽农938、辽星6RM121911230-0.56104铁路稻、辽盐糯、八里大穗、辽星13、公交13SNC⁃18RM2414240-1.1814岫岩不服劲、八里大穗、当地北海道、白刃、辽盐283250-1.6639元子二号、大红毛、改良北海道、太南稻、金早260-1.8229辽星6、辽星13、辽盐16、沈农129、红毛稻子RM28611160-2.8165岫岩不服劲、辽盐糯、八里大穗、丹粳8、开粳3号170-7.0310万大陆、老光头、老光头83、有芒紫叶稻、阿利西恩RM4714165-0.19144辽盐糯、八里大穗、丹粳8、开粳3号、辽农938SKC⁃18RM286111453.879岫岩不服劲、长春无芒、小白粳子桦甸白、丰产9号、123461602.3165丹粳8、开粳3号、辽农938、辽星6、辽盐1617013.0710有芒紫叶稻、丁旭稻、绢光、藤坂5号、陆奥穗秋波RM52762252.7221辽星6、辽盐16、辽粳912、二节稻、白大肚兴亚2300.3828铁路稻、辽盐糯、元子二号、红毛稻子、长白9号
表6(续)
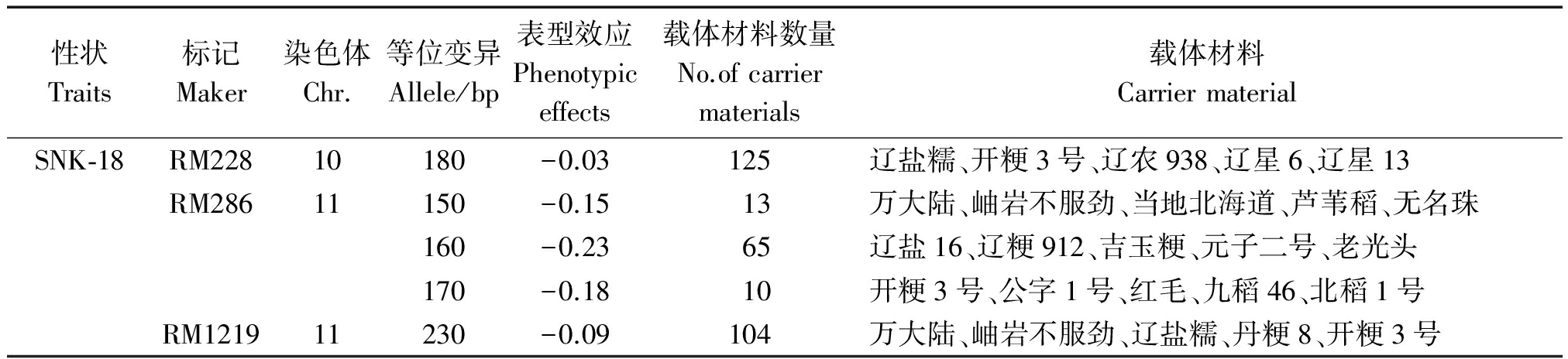
性状Traits标记Maker染色体Chr.等位变异Allele/bp表型效应Phenotypiceffects载体材料数量No.ofcarriermaterials载体材料CarriermaterialSNK⁃18RM22810180-0.03125辽盐糯、开粳3号、辽农938、辽星6、辽星13RM28611150-0.1513万大陆、岫岩不服劲、当地北海道、芦苇稻、无名珠160-0.2365辽盐16、辽粳912、吉玉粳、元子二号、老光头170-0.1810开粳3号、公字1号、红毛、九稻46、北稻1号RM121911230-0.09104万大陆、岫岩不服劲、辽盐糯、丹粳8、开粳3号
3 讨论与结论
关联分析是剖析复杂性状遗传基础的重要方法[33]。水稻的耐盐碱性是由多基因控制的数量性状[34]。寻找并控制水稻耐盐碱性的全部QTL,同时进行基因聚合对于提高水稻的耐盐碱性具有重要的意义。本研究所用的自然群体包括176个粳稻品种,这些品种具有广泛的地理分布,群体遗传变异较大。分别对13,18 d的取样结果分析可知,随处理时间增加,SNC、SKC和SNK的表型数值在平均数、变异范围等方面均有增加。同时,本试验与前人研究内容所不同的是,本试验针对水稻直播的种植方式,直接使用大庆地区标志性的中度苏打盐碱土进行胁迫处理,使研究更具有实际意义。
本研究利用176个粳稻品种组成的自然群体对SNC、SKC、SNK、SDG等4个耐盐碱相关性状进行了关联分析,在GLM和MLM 2种模型下共检测到了13个显著关联的位点,其中,有8个位点在2种模型下均被检测出显著关联。进一步利用这些位点挖掘到24个优异等位变异及相应的载体材料。为了提高育种效率,可以利用这些等位变异及其相应的载体材料进行粳稻芽期耐盐碱性状的改良,以期获得耐盐碱性优异的粳稻品种。
到目前为止,国内外在模拟盐碱条件下对水稻的耐盐碱性QTL定位进行了大量的研究[20,34-36]。本试验在2种模型条件下都检测到的与耐盐碱性状显著关联的8个位点中,7个为前人已经报道过的位点,1个为新发现的位点。通过比较相同标记和图谱,将本研究检测到的与耐盐碱性状相关联的位点与前人结果进行比较,发现位于第5染色体上与SDG-18显著关联的RM200与影响水稻耐盐指数的标记一致[24]。位于第4染色体上与SNC-13显著关联的RM471与Liang等[26]的研究结果中影响水稻死苗率的qDSRs4-2在同一染色体区段。位于第6染色体上的RM527与影响水稻地上部钾离子浓度的标记一致[27]。位于第1染色体上的RM1254与Wang等[28]检测到的与水稻种子发芽活力性状的标记一致。位于第4染色体上的RM241与Cheng等[30]研究结果中影响水稻在盐胁迫下存活率的QTL在相同染色体区段。RM1219在前人研究中未见报道。另外,本研究检测到的RM471、RM1219、RM527和RM286均与2个或2个以上性状显著关联。其中,RM1219与SNK-13、SNK-18和SDG均显著关联,同时RM1219也是在本研究中检测出的1个新位点,在前人研究中未见报道。很可能是与粳稻芽期耐盐碱性相关的新位点。关联分析能够有效、快速、准确地检测到与粳稻芽期耐盐碱性相关联的位点。本研究使用176份粳稻种质组成的自然群体,针对4个耐盐碱性状进行了粳稻芽期耐盐碱性鉴定,获得与粳稻芽期耐盐碱性显著关联的8个位点。同时挖掘出24个相关的优异等位基因及其相应的载体材料。可以利用以上研究制定适合黑龙江省盐碱地种植的粳稻品种。
参考文献:
[1] 安艳丽.水稻品种优选与田间管理[J].吉林农业,2017(12):48.
[2] 魏新彩. 基于HJ卫星影像的水稻种植面积遥感信息提取方法研究[D]. 武汉:湖北大学,2013.
[3] 梁银培,孙 健,索艺宁,等. 水稻耐盐性和耐碱性相关性状的QTL定位及环境互作分析[J]. 中国农业科学,2017,50(10):1747-1762.
[4] 于连鹏. 黑龙江省主栽水稻品种Pita,Pia和Piz-t抗瘟基因检测和抗性评价[D]. 大庆:黑龙江八一农垦大学,2017.
[5] 蔡永盛. 黑龙江盐碱地条件下不同产量类型水稻产量品质的比较研究[D]. 大庆:黑龙江八一农垦大学,2015.
[6] Famoso A N,Zhao K Y,Clark R T,et al. Genetic architecture of aluminum tolerance in rice(Oryza sativa) determined through Genome-Wide association analysis and QTL mapping[J]. PLoS Genetics,2011,7(8):747-757.
[7] Zheng H L,Liu B W,Zhao H W,et al. Identification of QTLs for salt tolerance at the germination and early seedling stage using linkage and association analysis in japonica rice[J]. Chinese Journal of Rice Science,2014,28(4):358-366.
[8] 王树国,张纪明. 水稻直播种植技术要点[J]. 农业与技术,2013(9):111-111.
[9] 付有权. 直播稻栽培技术探讨[J]. 经济技术协作信息,2014(14):79.
[10] 程海涛,苏 展,曹 萍,等. NaCl和Na2CO3胁迫对水稻籼粳杂交后代群体发芽与幼苗生育的影响[J]. 沈阳农业大学学报,2010,41(1):73-77.
[11] Remington D L,Thornsberry J M,Matsuoka Y,et al. Structure of linkage disequilibrium and phenotypic associations in the maize genome[J]. Proceedings of the National Academy of Sciences of the United States of America,2001,98(20):11479-11484.
[12] Weng J F,Xie C X,Hao Z F,et al. Genome-Wide association study identifies candidate genes that affect plant height in Chinese elite maize (Zea mays L.) inbred lines[J]. PLoS One,2011,6(12):e29229.
[13] Li Y,Huang Y,Bergelson J,et al. Association mapping of local climate-sensitive quantitative trait loci in Arabidopsis thaliana[J]. Proceedings of the National Academy of Sciences of the United States of America,2010,107(49):21199-21204.
[14] Huang X,Wei X,Sang T,et al. Genome-wide association studies of 14 agronomic traits in rice landraces[J]. Nature Genetics,2010,42(11):961-967.
[15] Niu Y,Xu Y,Liu X F,et al. Association mapping for seed size and shape traits in soybean cultivars[J]. Molecular Breeding,2013,31(4):785-794.
[16] 吴德志. 西藏野生大麦耐盐种质的发掘及其耐盐机制研究[D]. 杭州:浙江大学,2012.
[17] 印志同,杨庆华,倪正斌,等. 糯玉米芽苗期耐盐性鉴定及相关分子标记筛选[J]. 江苏农业学报,2012,28(2):278-283.
[18] 王建飞,陈宏友,杨庆利,等.盐胁迫浓度和胁迫时的温度对水稻耐盐性的影响[J].中国水稻科学,2004,18(5):75-80.
[19] Doyle J. Isolation of plant DNA from fresh tissue[J]. Focus,1991,12:13-15.
[20] 郑洪亮,刘博文,赵宏伟,等. 利用连锁和关联分析定位粳稻芽期及幼苗前期耐盐性 QTL[J]. 中国水稻科学,2014,28(4):358-366.
[21] 邢 军,常汇琳,王敬国,等. 盐,碱胁迫条件下粳稻Na+,K+浓度的QTL分析[J]. 中国农业科学,2015,48(3):604-612.
[22] Evanno G,Regnaut S,Goudet J. Detecting the number of clusters of individuals using the software STRUCTURE:a simulation study[J]. Molecular Ecology,2005,14(8):2611-2620.
[23] Bradbury P J,Zhang Z,Kroon D E,et al. TASSEL:software for association mapping of complex traits in diverse samples[J]. Bioinformatics,2007,23(19):2633-2635.
[24] Zhang T,Qian N,Zhu X,et al. Variations and transmission of QTL alleles for yield and fiber qualities in upland cotton cultivars developed in China[J]. PLoS One,2013,8(2):e57220.
[25] 聂新辉,尤春源,鲍 健,等. 基于关联分析的新陆早棉花品种农艺和纤维品质性状优异等位基因挖掘[J]. 中国农业科学,2015,48(15):2891-2910.
[26] Liang J L,Qu Y P,Yang C G,et al. Identification of QTLs associated with salt or alkaline tolerance at the seedling stage in rice under salt or alkaline stress[J]. Euphytica,2015,201(3):441-452.
[27] Zang J,Sun Y,Wang Y,et al. Dissection of genetic overlap of salt tolerance QTLs at the seedling and tillering stages using backcross introgression lines in rice[J]. Science in China Series C,2008,51(7):583-591.
[28] Wang Z F,Wang J F,Bao Y M,et al. Quantitative trait loci analysis for rice seed vigor during the germination stage[J]. Journal of Zhejiang University Science B,2010,11(12):958-964.
[29] Cui D,Wu D,Somarathna Y,et al. QTL mapping for salt tolerance based on snp markers at the seedling stage in maize(Zea mays L.)[J]. Euphytica,2015,203(2):273-283.
[30] Cheng L R,Wang Y,Meng L J,et al. Identification of salt-tolerant QTLs with strong genetic background effect using two sets of reciprocal introgression lines in rice[J]. Genome,2012,55(1):45-55.
[31] Cho Y G,Eun M Y,Mccouch S R,et al. The semidwarf gene,sd-1,of rice (Oryza sativa L.). Ⅱ. Molecular mapping and marker-assisted selection[J]. Theoretical & Applied Genetics,1994,89 (1) :54-59
[32] Bimpong I K,Manneh B,Diop B,et al. New quantitative trait loci for enhancing adaptation to salinity in rice from Hasawi,a Saudi landrace into three African cultivars at the reproductive stage[J]. Euphytica,2014,200(1):45-60.
[33] 董海娇. 基于全基因组关联分析的水稻分蘖角度和剑叶夹角的遗传基础解析[D]. 武汉:华中农业大学,2017.
[34] 潘世驹. 寒地水稻耐盐碱资源筛选及产量品质稳定性研究[D]. 大庆:黑龙江八一农垦大学,2016.
[35] 李 宁. 水稻苗期耐碱QTL分析[D]. 哈尔滨:东北农业大学,2016.
[36] Zheng T Q,Yang J,Zhong W G,et al. Novel loci for field resistance to black-streaked dwarf and stripe viruses identified in a set of reciprocal introgression lines of rice (Oryza sativa L.)[J]. Molecular Breeding,2012,29(4,SI):925-938.