小麦是世界上最重要的粮食作物之一,世界上有超过40%的人口以小麦为主食[1],黄淮麦区是中国最大的小麦主产区,在育种家的不断努力下,小麦品种的产量潜力还在不断地提高。从20世纪80年代中后期开始,小麦面粉的品质改良工作开始逐步受到育种家的重视,并已培育和推广了一批优质强筋的小麦品种[2]。小麦的面粉加工品质主要由其种子储藏蛋白控制,包括麦谷蛋白和麦醇溶蛋白,麦谷蛋白主要决定面团的弹性,是影响面筋质量的主要因素[3],由高分子量谷蛋白亚基(HMW-GS)和低分子量谷蛋白亚基(LMW-GS)组成。普通小麦的HMW-GS分别由位于第一同源群染色体长臂的Glu-A1、Glu-B1和Glu-D1位点的基因编码[4-6]。不同的HMW-GS对小麦品质的贡献并不相同,一般认为1、2*、7+9、14+15、17+18和5+10为优质亚基,具有较好的面粉加工品质[7-8]。LMW-GS分别由Glu-A3、Glu-B3 和Glu-D3位点的基因编码,较HMW-GS具有更丰富的多态性,LMW-GS通过和HMW-GS的互作效应,在一定程度上影响小麦的加工品质,优质的LMW-GS包括Glu-A3b、Glu-A3d、Glu-B3d和Glu-B3g位点[8]。目前,改良中国小麦面筋质量的主要途径是提高优质的HMW-GS和LMW-GS的频率[9-10],由于小麦LMW-GS的鉴定技术还不够十分完善,而小麦HMW-GS组成的鉴定技术较为成熟,因而,分析小麦HMW-GS组成已成为小麦品质育种亲本选配和后代选择的主要依据。2005年以来,针对黄淮麦区小麦新品种(系)的HMW-GS组成分析几乎每年都有报道[11-17],这为小麦品质育种的亲本选配提供了有价值的参考依据。然而,在现代育种过程中,由于长期的品种间杂交选育,尤其是对骨干亲本的重复利用,使现代小麦品种的遗传背景变得较为单一,其含有的HMW-GS组成结构,尤其是优质亚基的组成结构很难发生较大的改变。
本研究利用SDS-PAGE方法对221份黄淮麦区小麦新品系的HMW-GS组成进行鉴定,并对它们的系谱来源进行了分析,通过对近10年来已发表的黄淮麦区小麦品种(系)的HMW-GS分布频率进行比较,分析黄淮麦区小麦品种HMW-GS的组成结构变化,以期为小麦品质育种工作提供参考。
1 材料和方法
1.1 供试材料
所用材料为黄淮麦区2013-2014年冬水组和春水组参试的小麦新品系,共221份,由河南省农业科学院小麦研究中心赵虹研究员提供。
1.2 样品提取及SDS-PAGE电泳
参照Singh等[18]的方法提取高分子量谷蛋白及SDS-PAGE电泳分析。以中国春(Null, 7+8, 2+12)、豫麦34 (1, 7+8, 5+10)和济麦20 (1, 13+16, 4+12)为对照,按Payne和Lawrence[19]提出的命名系统命名亚基。
2 结果与分析
2.1 黄淮麦区小麦新品系的亚基组合及等位变异分析
利用SDS-PAGE技术对供试的221份黄淮地区小麦新品系材料的高分子量谷蛋白亚基组成进行了分析,其部分样品的HMW-GS电泳图谱见图1,各个亚基及亚基组合的等位变异及频率见表1。从表1中可以看出,供试材料共出现了13种亚基及亚基组合。在Glu-A1位点,1亚基出现频率最高,占供试材料的58.8%。在Glu-B1位点上,共出现7种亚基变异类型,其中出现频率最高亚基组合是7+9,占供试材料57.5%,强筋亚基组合7+8、17+18、14+15和13+16在供试材料中也有出现,分别占30.3%,2.3%,7.7%,0.9%。在Glu-D1位点上的变异类型有4种,其中强筋亚基组合5+10占供试材料的15.4%,另外,在以往普通小麦品种中很少出现的5+12亚基组合,占供试材料的9.0%,该亚基组合被认为是比5+10亚基组合更好的优质强筋亚基组合[20]。4+12亚基组合在本研究的供试材料中出现较大比例,达到31.7%。
2.2 黄淮麦区小麦新品系高分子量谷蛋白亚基的组合频率
对供试材料的高分子量谷蛋白亚基的组合频率进行了分析(表2),从表2中可以看出,供试材料的HMW-GS 组合类型比较丰富,有29种组合类型。其中出现频率最高的亚基组合形式是(Null, 7+9, 2+12),占全部供试材料的17.2%,其次为(1, 7+9,2+12)亚基组合,出现的频率为15.4%,表明黄淮麦区小麦新品系材料中弱筋的亚基组合类型仍然较多;(1,17+18,5+10)、(1,14+15,5+10)和(1,7+8,5+10)等强筋的亚基组合类型也有分布,但其出现频率相对较低,另外,还出现了(1,7+8,5+12)新的亚基组合类型,出现频率达到了3.6%。

1.中国春;2.豫麦34;3.济麦20;4.济麦06037;5.存麦17号;6.安农1208;7.保丰2018;8.濮兴5号;9.冠麦1号;10.航麦208;11.石11-4366;12.淮麦2230;13. SH4300;14.瑞泉麦168;15.豫圣麦19;16.邯生923;17.俊达109;18.瑞华055;19.现麦69;20.运旱20410。
1.Chinese spring;2. Yumai 34;3. Jimai 20;4. Jimai 06037;5. Cunmai 17;6. Annong 1208;7. Baofeng 2018;8. Puxing 5;9. Guanmai 1;10. Hangmai 208;11. Shi11-4366;12. Huaimai 2230;13. SH4300;14. Ruiquanmai 168;15. Yushengmai 19;16. Hansheng 923;17. Junda 109;18. Ruihua 055;19. Xianmai 69;20. Yunhan 20410.
图1 部分材料HMW-GS聚丙烯酰胺凝胶电泳图谱
Fig.1 The SDS-PAGE pattern of HMW-GS of partial test materials
表1 黄淮麦区小麦新品系HMW-GS等位基因变异及频率
Tab.1 Allelic variation and frequencies of HMW-GS of new wheat lines from Huang-Huai zone
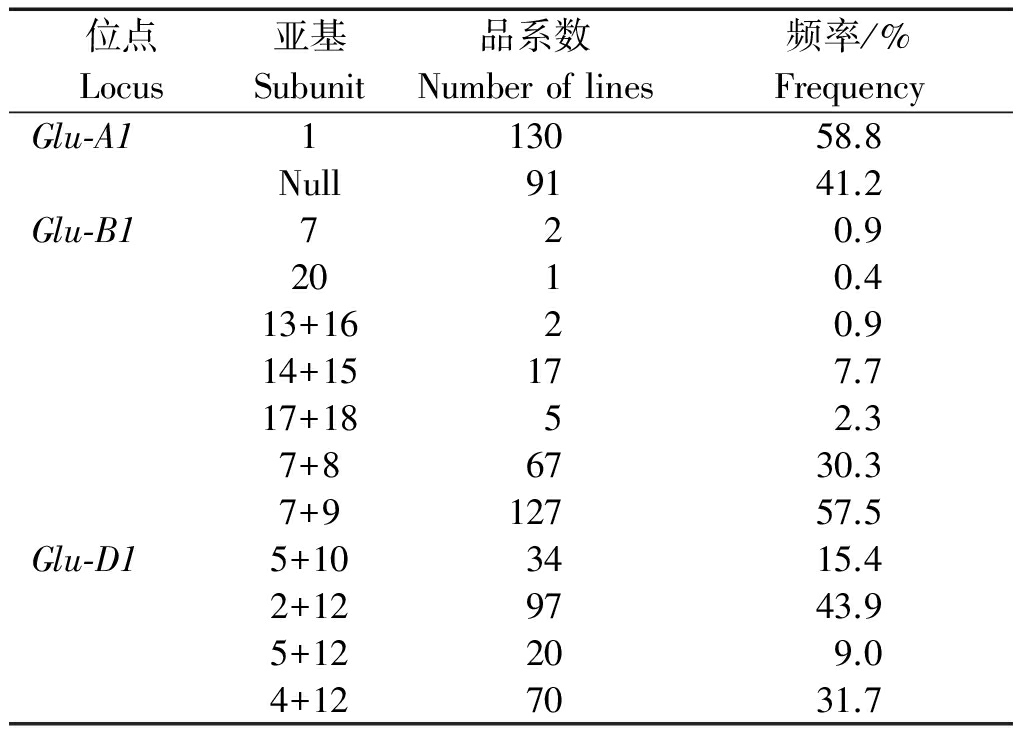
位点Locus亚基Subunit品系数Numberoflines频率/%FrequencyGlu⁃A1113058.8Null9141.2Glu⁃B1720.92010.413+1620.914+15177.717+1852.37+86730.37+912757.5Glu⁃D15+103415.42+129743.95+12209.04+127031.7
2.3 黄淮麦区小麦新品系的系谱来源分析
根据参加2013-2014年黄淮麦区的区试和预试材料提供的系谱来源进行分析,结果显示,直接以周麦16、矮抗58和周麦18为亲本培育的材料最多,分别有54,46和22个。另外,还有较多的直接以豫麦49和郑麦366为杂交亲本的材料。由于周8425B和豫麦2号是黄淮麦区的骨干亲本,利用这2个骨干亲本和其他小麦品种杂交选育出了一大批的小麦品种,由豫麦2号直接作为杂交亲本选育出周麦9号、新麦9号、豫麦34、豫麦49和石4185等品种,由周8425B和周麦9号作为杂交亲本选育出了大多数的周麦系列品种,在221份黄淮麦区小麦新品系中以周麦系列为直接杂交亲本的材料有95个,占到全部供试材料的43.0%。此外,郑麦366是由豫麦47和PH82-2-2杂交选育的,而豫麦47的杂交亲本之一为豫麦2号,矮抗58具有周8425B和豫麦49的血统。因而,在221份黄淮麦区小麦新品系中具有周8425B和豫麦2号血统的材料高达155个,占到全部供试材料的70.1%。以郑麦9023等小偃系列和鲁麦14等鲁麦系列为亲本的材料分别有34,40个,占全部供试材料的15.4%和18.1%。其中同时具有周8425B、豫麦2号和小偃系列血统以及周8425B、豫麦2号和鲁麦系列血统的材料均为20个,而同时具有小偃系列和鲁麦系列血统的材料仅有3个。其他的以豫麦18、冀麦系列、皖麦系列等为来源和未知来源的材料共35个。
表2 225份小麦新品系材料高分子量谷蛋白亚基组合类型
Tab.2 HMW-GS compositions of 225 new wheat lines

亚基组合类型Subunitcompositiontype品系数Numberoflines频率/%Frequency(Null,17+18,5+12)10.5(1,17+18,4+12)10.5(Null,14+15,5+12)10.5(1,20,2+12)10.5(Null,7+9,5+12)10.5(Null,17+18,4+12)10.5(1,17+18,5+10)20.9(1,13+16,4+12)20.9(1,14+15,5+10)20.9(1,14+15,4+12)20.9(1,7,5+10)20.9(1,14+15,2+12)31.3(Null,7+8,5+12)31.3(Null,14+15,4+12)41.8(Null,14+15,2+12)52.3(Null,7+8,5+10)52.3(1,7+8,2+12)52.3(Null,7+9,5+10)62.7(1,7+9,5+12)62.7(Null,7+8,4+12)62.7(1,7+9,5+10)83.6(1,7+8,5+12)83.6(Null,7+9,4+12)94.0(1,7+8,5+10)94.0(Null,7+8,2+12)115.0(1,7+8,4+12)209.0(1,7+9,4+12)2511.3(1,7+9,2+12)3415.4(Null,7+9,2+12)3817.2
2.4 黄淮麦区小麦品种(系)高分子量谷蛋白亚基组成的结构变化
根据已发表的黄淮麦区小麦新品种(系)高分子量谷蛋白亚基组成分析的文献[11-17],对近10年来的黄淮麦区小麦新品种(系)出现的HMW-GS频率进行了比较(表3),并分析了其HMW-GS的结构变化(图2)。由图2可以看出,在Glu-A1位点(图2-A),主要存在2种亚基类型,即1亚基和Null,自2008年以来,1亚基的出现频率高于Null类型,但二者的出现频率趋于接近, 2*亚基的出现频率相对较低。在Glu-B1位点(图2-B),存在的亚基类型较多,但主要的亚基组合类型是7+8和7+9,从2007年以来,7+9亚基组合类型出现的频率最高,并且一直维持较高的出现频率;另外,近10年来,对亚基组合14+15的利用从未停止,该组合具有类似 5+10 亚基的功能,与优质的面包加工品质呈正相关[21-22],但其出现频率相对较低,同时,在黄淮麦区小麦品种(系)中还出现了其他的优质强筋亚基组合类型,如13+16和17+18,其出现频率低,其中亚基组合17+18的亲本来源主要为烟农19。对于Glu-D1位点(图2-C),一直以来主要存在2+12和5+10 2种亚基组合类型,并且2+12亚基组合的出现频率高于5+10亚基,有趣的是,自2007年以来,2种亚基组合的出现频率整体呈平行的趋势,这表明育种工作者对2种亚基的利用趋于稳定,其主要的变化趋势是由于新出现的其他亚基组合(4+12和5+12)造成的,4+12亚基出现在2008,2010和2014*年,其出现频率分别为33.6%,21.3%和31.7%。
表3 近10年黄淮麦区小麦品种(系)高分子量谷蛋白亚基的出现频率
Tab.3 The occurrence frequencies of HMW-GS in the wheat varieties (lines) from Huang-Huai zone during the last decade
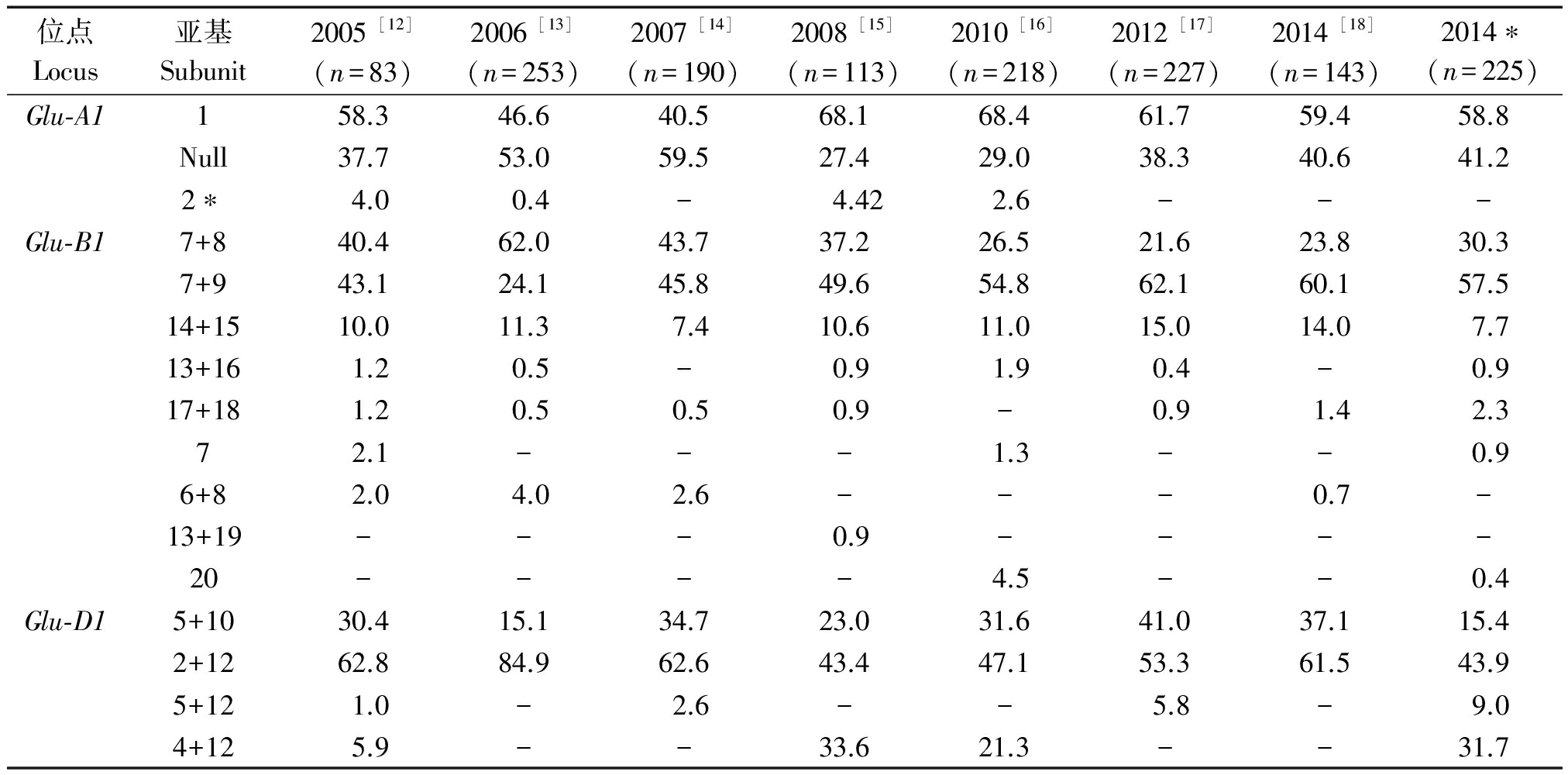
位点Locus亚基Subunit2005[12](n=83)2006[13](n=253)2007[14](n=190)2008[15](n=113)2010[16](n=218)2012[17](n=227)2014[18](n=143)2014∗(n=225)Glu⁃A1158.346.640.568.168.461.759.458.8Null37.753.059.527.429.038.340.641.22∗4.00.4-4.422.6---Glu⁃B17+840.462.043.737.226.521.623.830.37+943.124.145.849.654.862.160.157.514+1510.011.37.410.611.015.014.07.713+161.20.5-0.91.90.4-0.917+181.20.50.50.9-0.91.42.372.1---1.3--0.96+82.04.02.6---0.7-13+19---0.9----20----4.5--0.4Glu⁃D15+1030.415.134.723.031.641.037.115.42+1262.884.962.643.447.153.361.543.95+121.0-2.6--5.8-9.04+125.9--33.621.3--31.7
注:n.试验材料数; 2014*.本试验检测结果;-.表示数据不存在。
Note:n.The number of test materials; 2014*.The detected results in this work;-.The data is not found.
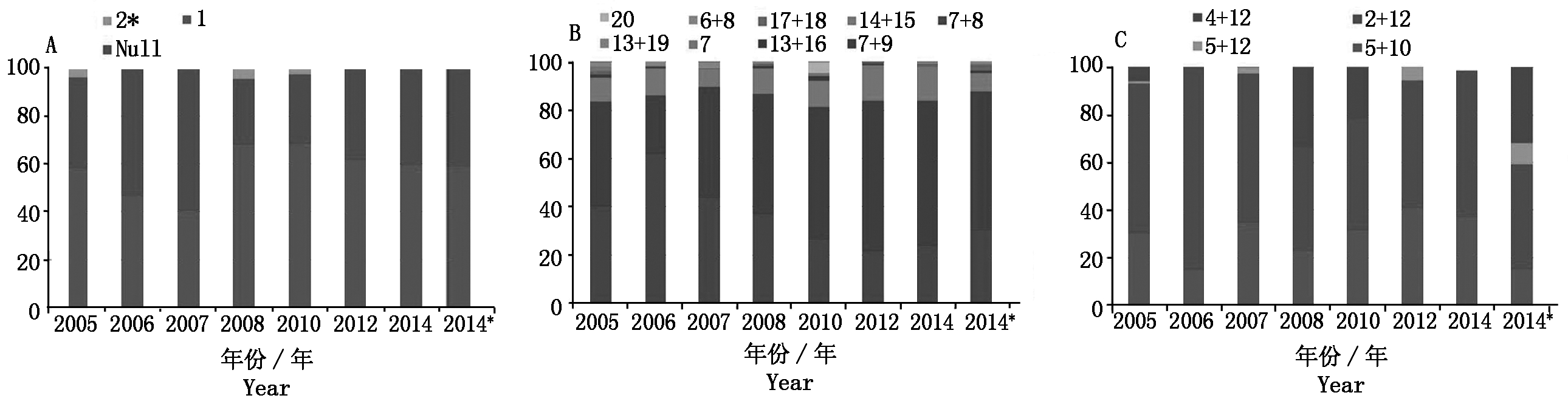
A.Glu-A1位点的结构变化;B.Glu-B1位点的结构变化;C.Glu-D1位点的结构变化。
A.The structure developments of Glu-A1 locus;B.The structure developments of Glu-B1 locus;C.The structure developments of Glu-D1 locus.
图2 近10年来黄淮麦区小麦品种(系)高分子量谷蛋白亚基组成的结构变化
Fig.2 The distribution trends of HMW-GS in the wheat varieties (lines)from Huang-Huai zone during the last decade
3 讨论与结论
长期以来,中国小麦育种主要以提高产量和改良抗病性为主,而小麦品种的面筋强度普遍较弱,造成其烘烤品质较差[23-25]。通过对20世纪90年代初的主栽品种的高分子量谷蛋白亚基组成进行分析,发现优质强筋的亚基频率低是中国小麦面筋强度弱的主要因素[26-27]。因而,选用具有不同HMW-GS的亲本杂交, 同时采用SDS-沉降值法和SDS-PAGE 凝胶电泳技术对其后代进行选择, 使不同基因位点上的优质强筋亚基聚合在一起, 以实现小麦籽粒蛋白质品质的逐步改良,是中国小麦品质育种的重要目标[28]。发现和鉴定优质强筋的亚基组合是小麦品质育种工作的基础,已知的Glu-B1位点上的13+16,17+18等亚基组合和Glu-D1位点上的5+10亚基组合都是较好的强筋亚基组合,5+12亚基组合最早是由![]() 等[29]在硬粒小麦与节节麦合成的六倍体小麦材料中发现的,研究表明,当合成的六倍体种中含有5+12亚基组合时,其烘烤品质和面包体积明显优于Glu-D1位点的其他亚基组合,如5+10和2+12亚基组合;Wang等[20]利用硬粒小麦×普通小麦的种间杂交获得了具有5+12亚基组合的小麦新种质,发现含有5+12亚基组合的小麦新种质在沉降值、湿面筋含量、面团形成时间和稳定时间等品质指标上显著优于其具有优质亚基组合5+10的姊妹系。5+12亚基组合在黄淮麦区小麦品种(系)中出现在2005,2007,2012,2014*年,其出现频率分别为1.0%,2.6%,5.8%和9.0%,在本研究中,共检测到8个小麦新品系含有1,7+8,5+12的亚基组合类型。由于目前对5+12亚基组合的研究较少,其亚基来源及其品质效应还需要进一步的分析。
等[29]在硬粒小麦与节节麦合成的六倍体小麦材料中发现的,研究表明,当合成的六倍体种中含有5+12亚基组合时,其烘烤品质和面包体积明显优于Glu-D1位点的其他亚基组合,如5+10和2+12亚基组合;Wang等[20]利用硬粒小麦×普通小麦的种间杂交获得了具有5+12亚基组合的小麦新种质,发现含有5+12亚基组合的小麦新种质在沉降值、湿面筋含量、面团形成时间和稳定时间等品质指标上显著优于其具有优质亚基组合5+10的姊妹系。5+12亚基组合在黄淮麦区小麦品种(系)中出现在2005,2007,2012,2014*年,其出现频率分别为1.0%,2.6%,5.8%和9.0%,在本研究中,共检测到8个小麦新品系含有1,7+8,5+12的亚基组合类型。由于目前对5+12亚基组合的研究较少,其亚基来源及其品质效应还需要进一步的分析。
不同位点上的高分子量谷蛋白亚基对小麦加工品质的影响不一样,通过对中国500 多份冬春小麦品种(系)的HMW-GS、LMW-GS与其面团流变学特性和面包面条品质的关系进行分析,发现HMW-GS和LMW-GS对小麦加工品质的影响顺序为Glu-D1>Glu-A3>Glu-B3>Glu-A1>Glu-B1>Glu-D3[30]。即Glu-D1位点对小麦加工品质的影响最大,因而,对Glu-D1位点上HMW-GS的改良应是中国小麦品质育种工作的重点,通过比较Glu-D1位点上的亚基组合出现频率的趋势可以看出,优质强筋的亚基组合5+10的出现频率始终低于2+12,自2007年5+10亚基组合的出现频率较2006年有所提高后,2007年直到现在(2014*年),黄淮麦区的小麦品种(系)Glu-D1位点上的5+10和2+12的出现频率呈平行趋势,2+12亚基组合的出现频率始终高于5+10亚基组合,其主要原因是选用的杂交亲本较为单一,主要的亲本来源均可追溯到周8425B和豫麦2号。在本研究中,Glu-D1位点上的4+12亚基的出现频率较高,有研究表明,我国的4+12亚基对面包品质的作用明显高于国外品种,在其亚基含量和HMW-GS总量均较高的情况下,其对面团品质的影响甚至高于5+10等亚基组合[31]。在对Glu-B1位点HMW-GS的改良上,不仅优质强筋的亚基组合17+18的利用频率较低,并且使用的杂交亲本来源也较为单一,其主要的亲本来源为烟农19。有研究表明,14+15亚基组合比7+8和7+9亚基组合对面包品质有更显著的正相关作用,亚基效应可表示为14+15>7+8>7+9[31]。另外,在含有公认的优质亚基组合5+10的品系中引入14+15,其品质评分也大大优于引入的其他组合[32]。目前,对于我国小麦品种的14+15亚基组合的鉴定和来源还存在争议,张学勇等[22]认为14+15亚基主要分布于冬麦区如黄淮冬麦区、长江中下游冬麦区和西南冬麦区,其亚基来源为St2422/464,并且在骨干亲本小偃6号和豫麦7号中均有该亚基组合。另外,利用SDS-PAGE技术还对中国小麦品种的HMW-GS类型进行了分类,发现14+15与20x+20y属于2种不同的亚基类型。而Gao等[33]分别利用SDS-PAGE、RP-HPLC、HPCE和MALDI-TOF-MS技术对小偃6号等小麦品种的Glu-B1位点进行鉴定,认为之前在我国小麦品种中发现的14+15与四倍体小麦Bidi17的20x+20y亚基带型一致,真正的14+15仅出现在一些欧洲的小麦品种里,而在亚洲国家中很少出现。在本研究中,黄淮麦区小麦新品种(系)的14+15亚基组合的带型与小偃6号一致,其出现频率较低。
近10年来黄淮麦区小麦品种(系)优质强筋的HMW-GS组成结构变化不明显,在Glu-B1位点上出现的17+18和13+16亚基组合频率较低,Glu-D1位点上2+12亚基组合的出现频率始终高于5+10亚基组合。其原因之一是其杂交亲本来源较为单一,大部分材料的亲本来源为周8425B、豫麦2号、小偃系列和鲁麦系列。
参考文献:
[1] Brenchley R, Spannag M, Pfeifer M, et al. Analysis of the bread wheat genome using whole-genome shotgun sequencing[J]. Nature, 2012, 491(7426): 705-710.
[2] 关二旗, 魏益民, 张 波, 等. 黄淮冬麦区部分区域小麦品种构成及品质性状分析[J]. 中国农业科学, 2012, 45(6): 1159-1168.
[3] Shewry P R.Wheat[J].J Exp Bot,2009,60(6):1537-1553.
[4] Payne P I, Corfield K G, Holt L M, et al. Correlation between the heritance of certain high-molecular-weight subunit of glutenin and bread-making quality in progenies of six crosses of bread wheat[J]. J Sci Food Agric, 1981, 32: 51-60.
[5] Payne P I. Genetics of wheat storage protein and effect of allelic variation on bread-making quality[J]. Ann New York Acad Sci, 1987, 38: 141-153.
[6] Zhang D L, He T T, Liang H H, et al. Flour quality and related molecular characterization of high molecular weight glutenin subunit genes from wild emmer wheat accession TD-256[J]. Journal of Agricultural and Food Chemistry, 2016, 64(24): 5128-5136.
[7] Tabiki T, Ikeguchi S, Ikeda T M. Effects of high-molecular-weight and low-molecular-weight glutenin subunit alleles on common wheat flour quality[J]. Breeding Science, 2006, 56(2): 131-136.
[8] Zhang D, Yuan Y, Su Y, Li S. Analysis of dough rheological property and gluten quality characteristics in wild emmer wheat(Triticum dicoccoides(Körn. ex Asch. et Graebn.) Schweinf.)[J]. Genetic Resources and Crop Evolution, 2016, 63: 675-683.
[9] 陈东升, 刘 丽, 董建力, 等. HMW-GS和LMW-GS及1BL/1RS对小麦品质性状的影响[J]. 作物学报, 2005, 31(4): 414-419.
[10] Guo X H, Hu J L, Wu B H,et al.Special HMW-GSs and their genes of Triticum turgidum subsp. dicoccoides accession D141 and the potential utilization in common wheat[J].Genetic Resources and Crop Evolution, 2016, 63(5):833-844.
[11] 宋亚珍, 王军卫, 侯文胜, 等. 黄淮流域小麦品种高分子量谷蛋白亚基遗传变异分析[J]. 西北农林科技大学学报:自然科学版, 2005, 33(6): 13-17.
[12] 张瑞奇, 胡 琳, 许为钢, 等. 黄淮冬麦区不同时期大面积推广品种的高分子量麦谷蛋白亚基组成分析[J]. 麦类作物学报, 2006, 26(2): 63-67.
[13] 牛吉山, 梁清志, 王保勤, 等. 黄淮麦区部分小麦种质资源高分子量谷蛋白亚基组成分析[J]. 麦类作物学报, 2007, 27(2): 245-249.
[14] 张维瑞, 纪利坤, 袁王俊, 等. 黄淮麦区小麦新品种(系)高分子量谷蛋白亚基组成分析[J]. 河南农业大学学报, 2008, 42(1): 6-10.
[15] 王美芳, 雷振生, 张学斌, 等. 黄淮冬麦区小麦品种(系)品质遗传组成及其效应分析[J]. 河南农业科学, 2010,39(10): 8-13.
[16] 张丽琴, 刘春雷, 杨 雪, 等. 黄淮麦区小麦新品种(系)高分子量谷蛋白亚基多态性分析[J]. 麦类作物学报, 2012, 32(1): 79-82.
[17] 麻珊珊, 刘 红, 武 军, 等. 黄淮麦区小麦新品种(系)HMW-GS组成分析[J]. 陕西农业科学, 2014, 60(11): 16-18,21.
[18] Singh N K, Shepherd K W, Cornish G B. A simplified of SDS-PAGE procedure for separating LMW subunits of glutenin[J]. J Cereal Sci,1991,14(3): 203-208.
[19] Payne P I, Lawrence G J. Catalogue of alleles for the complex gene loci, Glu-A1, Glu-B1, Glu-D1 which code for high molecular weight subunit of glutenin in hexaploid wheat[J]. Cereal Research Commun, 1983, 11: 29-35.
[20] Wang T, Li Z L, Ren Z L. Identification and characterization of a novel wheat line containing high molecular glutenin subunits 5+12[J]. Acta Agronomica Sinica, 2004,30(6): 544-547.
[21] 宋建民, 吴祥云, 刘建军, 等. 小麦品质的麦谷蛋白亚基评定标准研究[J]. 作物学报, 2003, 29(6): 829-834.
[22] 张学勇, 庞斌双, 游光霞, 等. 中国小麦品种资源Glu-1位点组成概况及遗传多样性分析[J]. 中国农业科学, 2002,35(11): 1302-1310.
[23] 齐琳娟, 胡学旭, 周桂英, 等. 2004-2011年中国主产省小麦蛋白质品质分析[J]. 中国农业科学, 2012, 45(20): 4242-4251.
[24] 张平平, 张 瑜, 王化敦, 等. 小麦Glu-B1x7沉默基因序列分析及对加工品质的影响[J]. 核农学报, 2018, 32(4): 708-714.
[25] Wang H, Zhang H, Li B, et al. Molecular cytogenetic characterization of new wheat-Dasypyrum breviaristatum introgression lines for improving grain quality of wheat[J]. Frontiers in Plant Sciences, 2018, 9:365.
[26] Liu L, He Z H, Yan J, et al. Allelic variation at the Glu-1 and Glu-3 loci, presence of the 1B.1R translocation, and their effects on mixographic properties in Chinese bread wheats[J]. Euphytica, 2005, 142(3):197-204.
[27] Filip E. Composition of high molecular weight glutenin subunits in polish common wheat cultivars (Triticum aestivum L.)[J]. Journal of Food Quality, 2018(3):1-8.
[28] 何中虎, 晏月明, 庄巧生, 等. 中国小麦品种品质评价体系建立与分子改良技术研究[J]. 中国农业科学, 2006, 39(6):1091-1101.
[29] ![]() R J, Zarco-Hernandez J, Mujeeb-Kazi A. Glutenin subunit compositions and bread-making quality characteristics of synthetic hexaploid wheat derived from Triticum turgidum×Triticum tauschii(coss.) Schmal Crosses[J]. J Cereal Sci, 1995,21(1): 15-23.
R J, Zarco-Hernandez J, Mujeeb-Kazi A. Glutenin subunit compositions and bread-making quality characteristics of synthetic hexaploid wheat derived from Triticum turgidum×Triticum tauschii(coss.) Schmal Crosses[J]. J Cereal Sci, 1995,21(1): 15-23.
[30] He Z H, Liu L, Xia X C, et al. Composition of HMW and LMW glutenin subunits and their effects on dough properties, Pan bread, and noodle quality of Chinese bread wheats[J]. Cereal Chemistry, 2005, 82(4): 345-350.
[31] 高 翔, 雷 玲, 董 剑, 等. 小麦高分子量谷蛋白亚基效应的比较研究[J]. 西北植物学报, 2005, 25(12): 2443-2446.
[32] Deng Z Y, Tian J C, Sun G X. Influence of high molecular weight glutenin subunit substitution on rheological behaviour and bread-baking quality of near-isogenic lines developed from Chinese wheats[J]. Plant Breeding, 2005, 124(5): 428-431.
[33] Gao L Y, Ma W J, Chen J, et al. Characterization and comparative analysis of wheat high molecular weight glutenin subunits by SDS-PAGE, RP-HPLC, HPCE, and MALDI-TOF-MS[J]. Journal of Agricultural and Food Chemistry, 2010, 58(5): 2777-2786.