油菜是我国重要的油料和经济作物之一,菜籽油是我国主要植物食用油之一。菜籽油不仅富含人体所必需的氨基酸和脂肪酸,而且随着脂肪酸含量的提升增加了植物油的化学稳定性,能较长时间地储存[1]。因此,脂肪酸的含量决定了菜籽油的品质。酰基载体蛋白(Acyl carrier protein,ACP)参与脂肪酸和甲羟戊酸的合成及脂肪酸的脱氢反应[2],在脂肪酸合成的过程中酰基载体蛋白是一个最为关键的蛋白质[3],该蛋白是一个具有保守丝氨酸残基的小分子量的可溶酸性蛋白质,其辅基4′-磷酸泛酰巯基乙胺与ACP中的丝氨酸残基通过磷酸酯键相连[4-5],另一端的-SH自由基通过硫酯键连接脂酰基[6],其在酶反应过程中起着运输的作用[7-9],将脂酰基从一个酶反应转移到另一个酶反应[10]。有科学家指出,在拟南芥中有多种不同的质体ACP,其中包括5种质体ACP以及3种线粒体ACP[11],这些ACP有不同的调控方式和表达模式[12-13]。研究发现,ACP异构体的种类及差异表达与植物脂肪酸中不饱和脂肪酸组成及占总脂肪酸的比例有密切关系[4,14-15];植物ACP的表达方式共有组成型表达和组织特异性表达及细胞特异性表达3种不同的方式[12,16]。目前,植物中ACP基因的研究进展十分迅速,研究发现,将ACP基因转入芸薹属作物中,在叶中表达量增加数倍,表达量增加的同时16∶3的脂肪酸明显减少,亚麻酸(18∶3)的含量明显增加,种子中油酸和亚油酸的含量明显增加[17],但是在种子内总脂肪酸含量没有明显变化[4]。因此,改变ACP基因的表达有可能改变菜籽油中脂肪酸的组成及含量[8]。然而,现在对ACP5基因的研究报道还不多见,其生物信息学分析很不完善及其在种子中的表达分析尚未见到报道。本研究对BnACP5基因克隆,并进行生物信息学分析,预测其结构与功能,旨在为高油酸油菜品种选育提供理论基础。
1 材料和方法
1.1 试验材料
湘油15由湖南农业大学油料改良中心提供。RNA Kit、 反转录、荧光定量、琼脂糖凝胶纯化回收试剂盒均在北京全式金生物技术有限公司购买;PrimeSTAR GXL DNA Polymerase、pMD-19T Vector Cloning kit等从TaKaRa公司购买。
1.2 试验方法
1.2.1 RNA提取及cDNA合成 参照 RNA Kit试剂盒说明书分别提取各时期种子的总RNA,所提RNA用1.5%琼脂糖凝胶电泳检测其完整性,用Nanodrop 2000检测RNA的纯度和浓度,于-80 ℃保存。用反转录试剂盒参照说明书反转录合成cDNA第一链及去除gDNA,于-20 ℃保存[18]。
1.2.2 试验引物设计及基因克隆 从TAIR数据库得到拟南芥AtACP5基因编号(AT5G27200),并将该基因编号于Brassica Database数据库(http://brassicadb.org/brad/index.php)Blast得到一段序列,利用Primer Premier 5软件在这段序列两端设计一对引物ACP5-F/ACP5-R(表1),以30 d种子的cDNA为模板,用引物ACP5-F(CCGCCATCTCTCTCT
CTTGATC)/ACP5-R(GAACAGAGGCACATTTAAGC
GG)进行PCR扩增。PCR反应体系:模板1 μL,dNTPs 4 μL,5×Buffer 10 μL,ACP5-F/ACP5-R各1 μL, DNA聚合酶1 μL。反应程序:95 ℃预变性5 min;95 ℃变性50 s,52 ℃复性1 min,72 ℃延伸1 min,设35个循环;72 ℃延伸10 min。PCR产物在1.5%琼脂糖凝胶电泳检测,将检测到的目的条带用琼脂糖凝胶纯化回收试剂盒回收,将回收产物连接到pMD-19T载体后转化大肠杆菌,并测序。
1.2.3 甘蓝型油菜BnACP5基因的序列对比及聚类分析、生物信息学分析 利用DNAMAN软件对BnACP5基因的序列进行对比;结合Mega 6.06软件、NCBI的CDD在线工具、ExPASy蛋白质分析系统中的ProtParam在线工具、ProtScale在线分析软件、在线软件Signalp 4.1 server、TMHMM Server v 2.0在线软件、SOPMA、Netphos 2.0 server、Psort Ⅱ Prediction分别对BnACP5基因进行聚类分析及其编码蛋白的结构域、基因编码蛋白的理化性质、疏水性/亲水性、信号肽、跨膜结构、二级结构、磷酸化位点、亚细胞定位分析及预测[19]。
1.2.4 甘蓝型油菜BnACP5基因相对表达量分析 以湘油15不同生长时期的种子的cDNA为模板,从已获得的BnACP5基因中设计荧光定量PCR引物QPCR-F(GACATCCCATCCCTGCTC)/QPCR-R(ATC
CCGAACTCTTCCTCC),从Brassica Database中得到甘蓝型油菜BnUBC21基因序列并设计一对内参引物UBC21-F(CCTCTGCAGCCTCCTCAAGT)/UBC21-R(TATCTCCCCTGTCTTGAAATGC)。具体操作参照荧光定量试剂盒的说明书进行,数据采用CFX Manager 软件中-ΔΔCT法进行分析。
2 结果与分析
2.1 BnACP5基因的克隆
以提取湘油15号30 d种子中的RNA为模板,通过RT-PCR反转录成cDNA第一条链,用引物ACP5-F/ACP5-R PCR进行PCR扩增,扩增出长度约为500 bp的基因片段(图1)。用琼脂糖凝胶纯化回收试剂盒将检测到的目的基因条带回收,将回收产物连接pMD-19T载体并转化大肠杆菌,经菌落PCR检测呈阳性,即获得阳性克隆。将其与Brassica Database比对的一段417 bp的CDs序列命名为BnACP5。
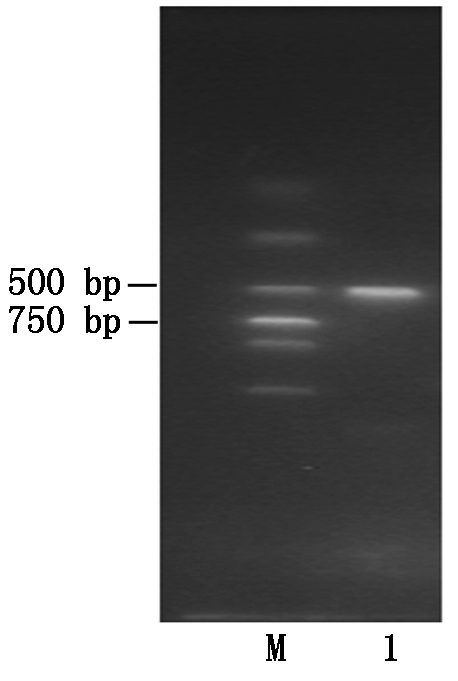
M.Trans2K DNA Markers;1.PCR产物。
M.Trans2K DNA Markers;1.PCR results.
图1 BnACP5 PCR产物电泳
Fig.1 The electrophoresis of BnACP5 PCR products
2.2 BnACP5蛋白生物信息学分析
2.2.1 甘蓝型油菜ACP5氨基酸序列分析 通过NCBI的CDD在线工具分析该蛋白的结构域,结果显示,该蛋白含有acyl_carrier结构域(57-133)。结构域中有磷酸泛酰巯基乙胺结合位点(Phosphopantetheine attachment site),该位点包含在保守的Asp-Ser-Leu (DSL)基序中,该基序为磷酸泛酞琉基乙胺基转移酶家族成员识别。该保守结构域属于磷酸泛酰巯基乙胺结合蛋白超家族(PP-binding superfamily)。通过NCBI对BnACP5进行BlastX比对,结果显示,BnACP5共编码138个氨基酸,BnACP5与白菜型油菜的ACP5序列相似性最高,达到98%。利用DNAMAN软件将BnACP5蛋白与拟南芥(NP_198072.1)、甘蓝(XP_013607498.1)、白菜型油菜(XP_009111933.1)的氨基酸序列进行比对(图2),结果发现,这4个物种的氨基酸序列中的acyl_carrier结构域具有高度的相似性;ACP氨基酸序列比对表明,植物ACP磷酸泛酰巯基乙胺结合位点周围的氨基酸序列高度保守。
2.2.2 甘蓝型油菜ACP5氨基酸聚类分析 为了分析BnACP5的进化发育,结合Mega 6.06软件将BnACP5的氨基酸序列与从NCBI的nr数据库中选取了9个相似性高的氨基酸序列:拟南芥(NP_198072.1)、白菜型油菜(XP_009111933.1)、甘蓝(XP_013607498.1)、山葵(XP_006394987.1)、亚麻荠(XP_010455149.1)、醉蝶花(XP_010524895.1)、玉山筷子芥(XP_002874402.1)、荠菜(XP_006289890.1)、高山南芥(KFK26465.1),进行UPGMA聚类分析,结果见图3。根据亲缘关系的远近可分为4类:甘蓝型油菜、白菜型油菜与甘蓝为一类,醉蝶花为一类,其他物种共为一类,其中,甘蓝型油菜与白菜型油菜亲缘关系最近。
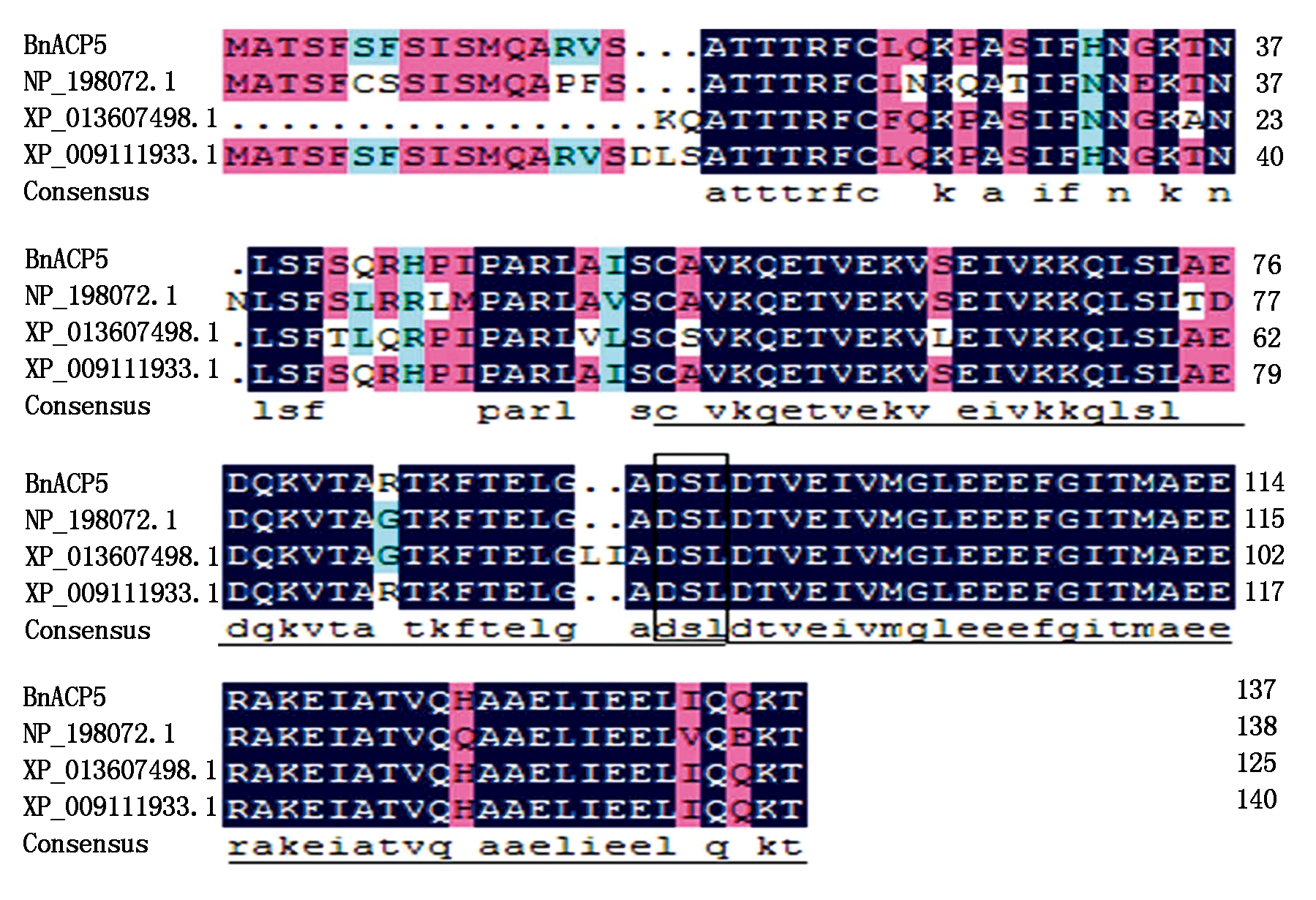
实线代表acyl_carrier结构域;方框代表DSL基序。
The acyl_carrier-domain marked with solid line;The DSL motif marked with box.
图 2 甘蓝型油菜BnACP5编码蛋白与其他物种蛋白的氨基酸序列对比
Fig.2 Homology analysis of the deduced amino acid sequences from BnACP5 and those of other plant species
2.2.3 甘蓝型油菜BnACP5氨基酸的理化性质分析 利用ProtParam分析BnACP5基因编码的蛋白的理化性质,结果显示,BnACP5由138个氨基酸组成,分子式为C667H1093N183O211S6,相对分子质量为15.25 ku,理论等电点(pI)为5.94,其不稳定参数为43.03,属于不稳定蛋白。该基因编码的多肽含丙氨酸(Ala,A)最多,占总氨基酸的11.6%;其次是谷氨酸(Glu,E),占总氨基酸的10.9%。其中,不含色氨酸(Trp,W)、酪氨酸(Tyr,Y)、甲硫胺酸(Pyl,O)、硒半胱氨酸(Sec,U),其总疏水平均系数(GRAVY)为-0.148,属亲水性蛋白。为了进一步说明BnACP5基因编码的蛋白是一个亲水蛋白,运用ProtScale软件进一步分析(图4),结果显示,基线上部曲线明显少于下部曲线,说明大部分氨基酸为亲水性氨基酸,这也说明该蛋白为亲水蛋白。
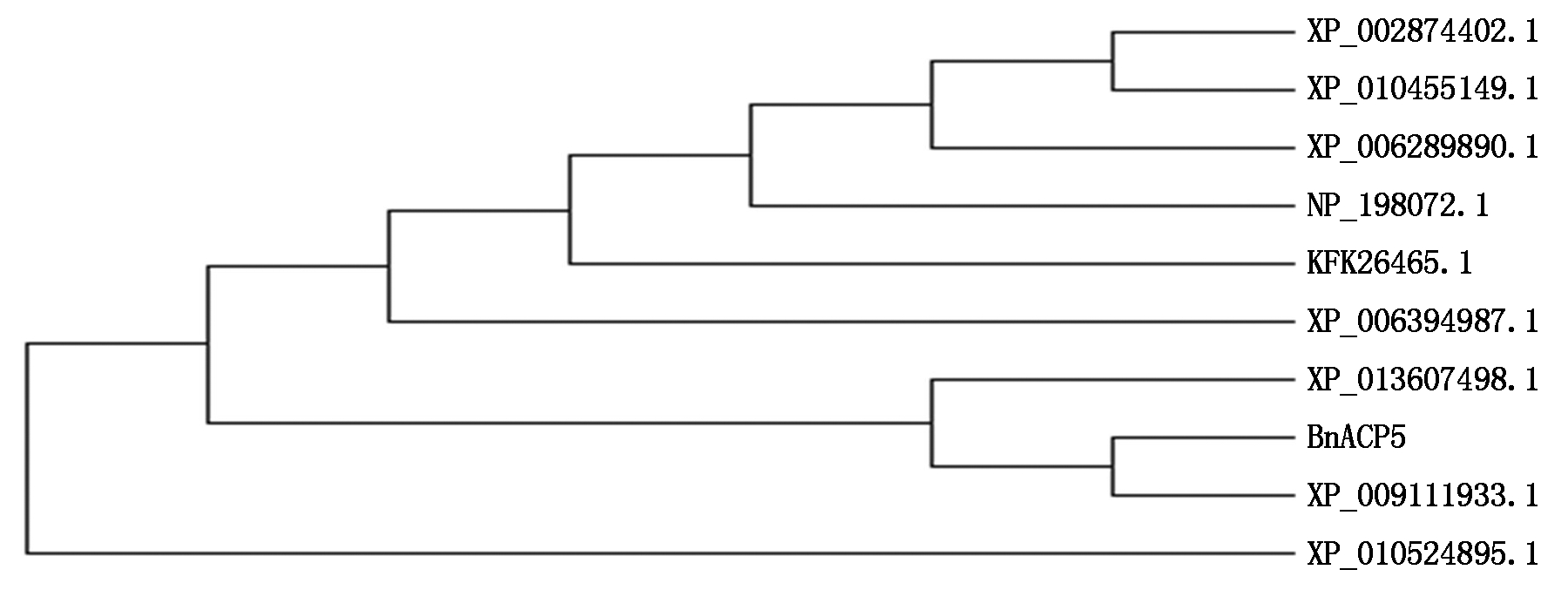
NP_198072.1.拟南芥;XP_009111933.1.白菜型油菜;XP_013607498.1.甘蓝;XP_006394987.1.山葵;XP_010455149.1.亚麻荠;XP_010524895.1.醉蝶花;XP_002874402.1.玉山筷子芥;XP_006289890.1.荠菜;KFK26465.1.高山南芥。
NP_198072.1.Arabidopsis thaliana; XP_009111933.1.Brassica rapa;XP_013607498.1.Brassica oleracea; XP_006394987.1.Wasabi japonica;XP_010455149.1.Camelina sativa L.Crantz. ; XP_010524895.1.Cleome spinosa L.; XP_002874402.1.Arabis lyrata L.; XP_006289890.1.Capsella bursa-pastoris; KFK26465.1.Arabis alpine.
图 3 甘蓝型油菜ACP5氨基酸与其他相似性高的氨基酸聚类
Fig.3 Phylogenetic tree of amino acid of ACP5 of B.napus and other similar amino acids
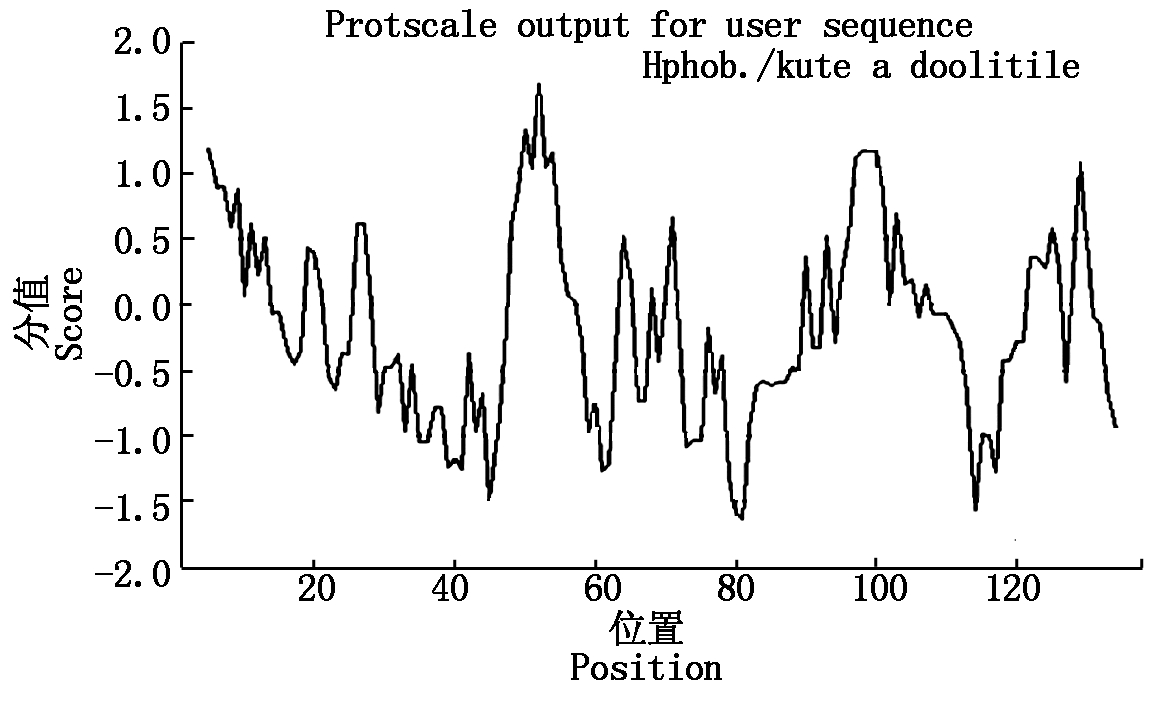
图4 BnACP5基因编码蛋白的亲疏水性预测
Fig.4 Hydrophilicity and hydrophobicity prediction of BnACP5 gene coding protein
2.2.4 甘蓝型油菜BnACP5信号肽及跨膜结构预测 在Signa1p 4.1在线软件上对其编码蛋白的信号肽进行预测(图5),结果显示,第18个氨基酸处为信号肽C和Y的最大切割点,分值分别为0.260和0.239,第4个氨基酸处为信号肽S的最大切割点,分值为0.305。由此可知,BnACP5无信号肽序列,属于非分泌蛋白。从TMHMM Server v 2.0对BnACP5跨膜结构进行预测,结果显示,该蛋白不含跨膜结构,为非跨膜蛋白。
2.2.5 甘蓝型油菜BnACP5基因编码蛋白的二级结构预测及磷酸化位点预测、亚细胞定位 利用SOPMA在线软件预测其二级结构,结果显示,该蛋白的二级结构类型为:α螺旋为59.42%、无规则卷曲为23.19%、延伸链为14.49%,β转角比例最小,为2.90%。运用Netphos 2.0 server在线软件对其编码蛋白进行磷酸化位点预测,结果显示(图6),该蛋白序列中含有13个苏氨酸(Threonine,T)和12个丝氨酸(Serine,S),其中18个可能是磷酸化位点。通过Psort Ⅱ Prediction预测该蛋白位于线粒体内。
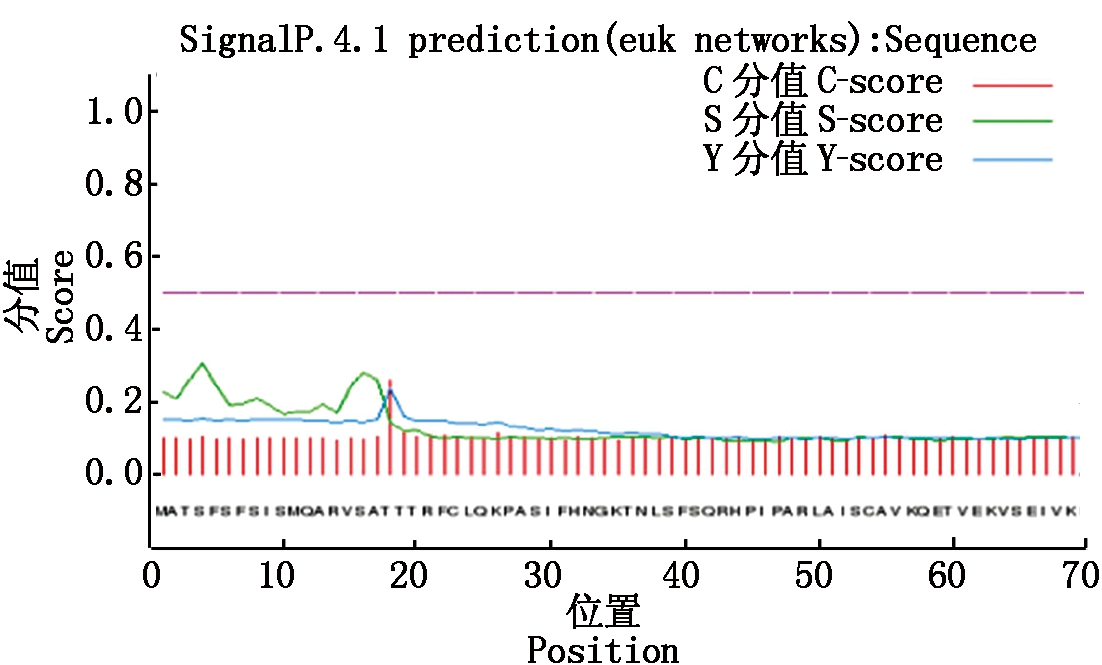
图5 BnACP5基因编码蛋白信号肽预测
Fig.5 Singal peptide prediction of BnACP5 gene coding protein
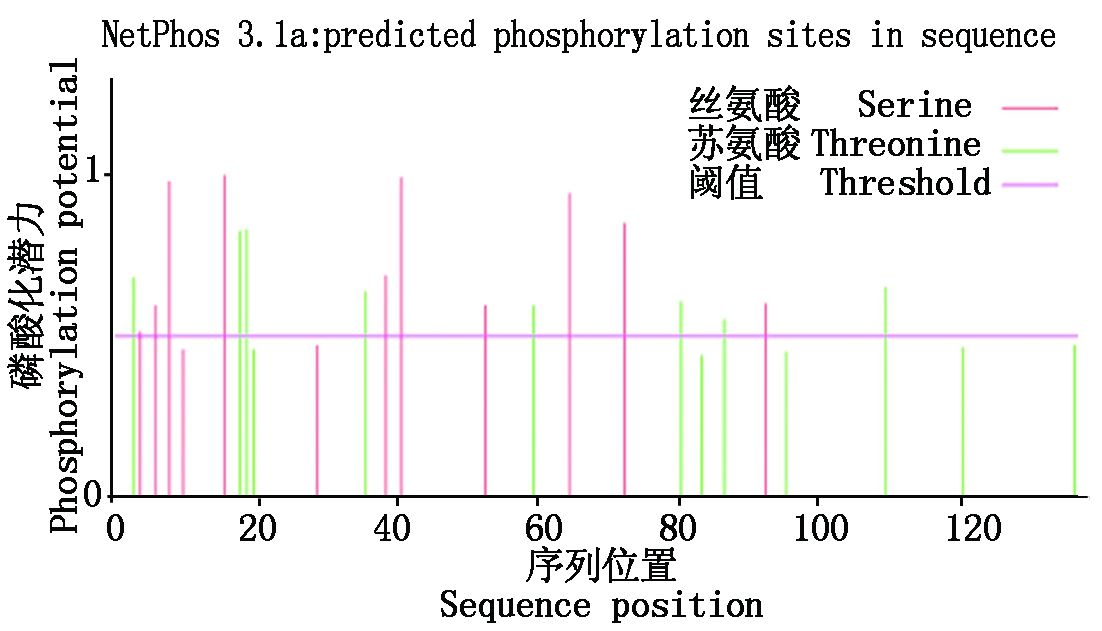
图6 BnACP5基因编码蛋白磷酸化位点预测
Fig.6 Phosphorylation site prediction of BnACP5 gene coding protein
2.3 BnACP5基因相对表达分析
由图7可知,该基因的表达量在种子发育的不同时期呈一定规律的变化,在授粉后15 d种子相对表达量就达到峰值;在20 d时,呈下降趋势;在25~35 d种子BnACP5基因的相对表达量呈上升趋势,之后迅速下降;在45 d,种子BnACP5基因的相对表达量已降到最低水平。
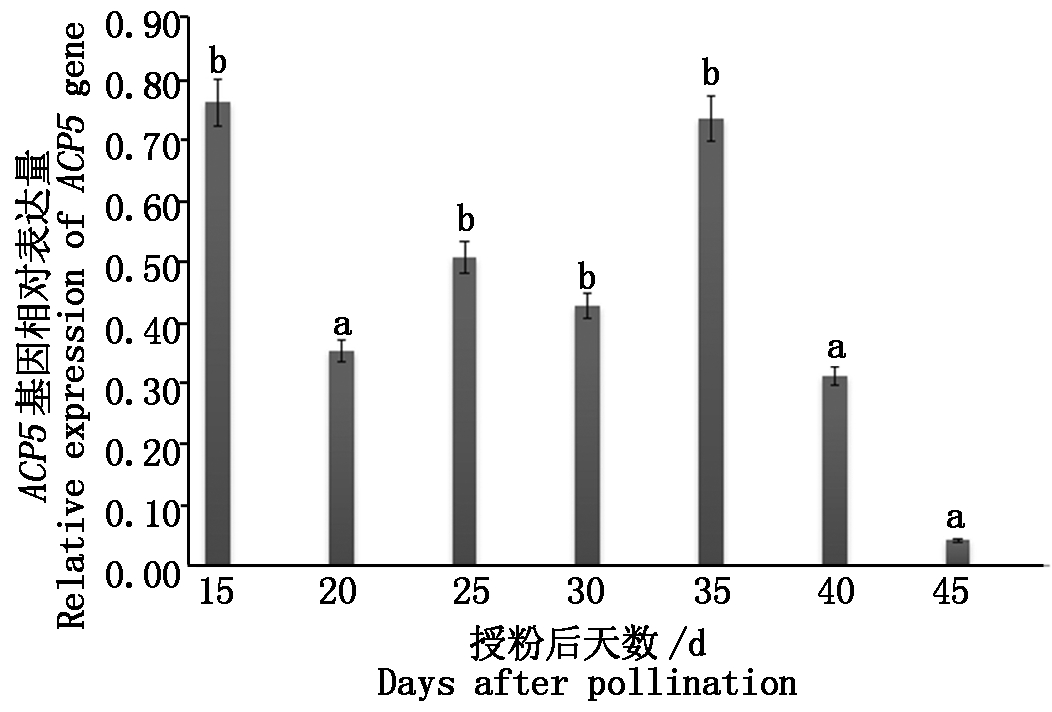
不同小写字母表示在0.05水平上差异显著。
Different small letters indicate significant at the 0.05 level.
图7 甘蓝型油菜种子不同时期中BnACP5基因的表达情况
Fig.7 The expression of BnACP5 gene at rapeseed seed different growth stages
3 讨论
对BnACP5蛋白的氨基酸序列分析结果显示,甘蓝型油菜与白菜型油菜的ACP5同源性最高,为98%,并且该蛋白具有高度保守的PP-binding结构域,也具有与丝氨酸残基共价结合的磷酸泛酸盐,是一个典型的酰基载体蛋白[20]。对该家族氨基酸UPGMA聚类分析结果显示,BnACP5与白菜型油菜、甘蓝亲缘关系较近,与其他植物亲缘关系较远,由此可推测,ACP5在进化中相对保守。对甘蓝型油菜BnACP5基因编码的蛋白理化性质分析显示,该蛋白为不稳定的亲水性蛋白,不稳定参数为43.03,是一个可溶的酸性蛋白,这些与Branen 等[14]研究结果基本吻合。研究发现,AtACP具有高度保守特性,对于脂肪酸的含量和组成具有重要的作用[3]。本研究在甘蓝型油菜克隆出一条BnACP5 CDs序列,对其蛋白亚细胞定位结果也显示其在线粒体上,这与拟南芥中的AtACP基因研究结果基本一致[11]。
近年来,大量的研究表明,ACP基因与脂肪酸合成有关,因此,研究其在种子的表达趋势,对其功能的挖掘有重要的意义。组织特异性表达分析表明,BnACP5基因在甘蓝型油菜种子发育过程中高度表达。由此表明,BnACP5基因可能与种子的发育有关。本研究中,BnACP5在15 d达到峰值,20 d下降,25~35 d呈上升趋势,此后迅速下降,45 d表达量降到最低水平。在拟南芥[4]和芥菜[17]中研究发现,ACP超表达改变了不饱和脂肪酸所占的比例,但提高了种子的油酸(18∶1)和亚油酸(18∶2)含量。高建芹[21]、李成磊[22]等研究表明,在甘蓝型油菜种子发育过程中的油酸(18∶1)与亚油酸(18∶2)呈极显著负相关。油酸(18∶1)的含量呈现先上升后下降的趋势;而亚油酸(18∶2)开花后15 d含量最高,随着角果的发育进程含量下降。验证了本试验前期BnACP5表达量高,后期总体呈下降趋势的结论。
本研究克隆了甘蓝型油菜BnACP5基因,对其表达模式进行了qPCR分析,这为后续开展功能验证打下了坚实的基础。
参考文献:
[1] 刘睿洋.甘蓝型油菜BnFAD2基因的结构及功能分析[D].长沙:湖南农业大学,2016.
[2] 单 雷,唐桂英,徐平丽,等.花生质体型酰基载体蛋白基因5′侧翼调控序列的克隆与分析[J].作物学报,2014,40(3):381-389.
[3] Nguyen C,Haushalter R W,Lee D J,et al.Trapping the dynamic acyl carrier protein in fatty acid biosynthesis[J].Nature,2014,505:427-431.
[4] 牟晶晶.小麦硬脂酰基载体蛋白脂肪酸去饱和酶基因(TaSSI2-1)的功能分析[D].泰安:山东农业大学,2015.
[5] 李志平,江贤章,田宝玉,等.裂殖壶菌酰基载体蛋白(ACP)基因的克隆与表达[J].微生物学通报,2008(2):256-260.
[6] 谭晓风,王威浩,刘卓明,等.油茶ACP基因的全长cDNA克隆及序列分析[J].中南林业科技大学学报,2008(4):8-14.
[7] 王树彦,韩 冰,周四敏,等.油用亚麻可溶性糖、脂肪含量与硬脂酰-酰基载体蛋白脱氢酶基因表达相关性分析[J].作物杂志,2016(4):56-61.
[8] 陶永佳,薛永常.酰基载体蛋白在不同代谢通路中的作用[J].生命的化学,2016,36(6):914-917.
[9] 牛 蓓,徐 莺,宋 君,等.乌桕硬脂酰-酰基载体蛋白脱饱和酶基因的克隆及表达[J].西南农业学报,2016,29(8):1806-1812.
[10] Akey D L,Razelun J R,Tehranisa J,et al.Crystal structures of dehydratase domains from the curacin polyketide biosynthetic pathway[J].Structure,2010,18(1):94-105.
[11] 李孟军,史占良,郭进考,等.植物酰基载体蛋白基因家族序列分析[J].华北农学报,2010,25(S):1-6.
[12] Bonaventure G,Ohlrogge J B.Differential regulation of mRNA levels of acyl carrier protein isoforms in Arabidopsis[J].Plant Physiology,2002,128(1):223-235.
[13] Hlousek-Radojci A,Post-Beittenmiller D,Ohlrogge J B.Expression of constitutive and tissue-specific acyl carrier protein isoforms in Arabidopsis[J].Plant Physiology,1992,98(1):206-214.
[14] Branen J K,Shintani D K,Engeseth N J.Expression of antisense acyl carrier protein-4 reduces lipid content in Arabidopsis leaf tissue[J].Plant Physiology,2003,132(2):748-756.
[15] Roughan G,Matsuo T.The influence of endogenous acyl-acyl carrier protein concentrations on fatty acid compositions of chloroplast glycerolipids[J].Archives of Biochemistry and Biophysics,1992,297(1):92-100.
[16] Aluru M R,Mazourek M,Landry L G,et al.Differential expression of fatty acid synthase genes,Acl,Fat and Kas,in Capsicum fruit[J].Journal of Experimental Botany,2003,54(388):1655-1664.
[17] Jha J K,Sinha S,Maiti M K,et al.Functional expression of an acyl carrier protein (ACP) from Azospirillum brasilense alters fatty acid profiles in Escherichia coli and Brassica juncea[J].Plant Physiology and Biochemistry,2007,45(6/7):490-500.
[18] 虢 慧,贺 慧,吴宁柔,等.不同关键酶基因在油菜种子发育进程中的表达[J].分子植物育种,2017,15(8):3030-3035.
[19] 邢 蔓,刘少锋,邬贤梦,等.甘蓝型油菜GPAT9基因克隆与生物信息学分析[J].分子植物育种,2016,14(12):3282-3288.
[20] 赵 艳,杨晓杰,范震宇,等.大豆酰基载体蛋白硫酯酶基因及其启动子表达方式的初步分析[J].大豆科学,2012,31(5):714-717.
[21] 高建芹,浦惠明,戚存扣,等.高含油量油菜种子和果皮油分积累及主要脂肪酸的动态变化[J].中国油料作物学报,2009,31(2):173-179.
[22] 李成磊,付三雄,戚存扣.甘蓝型油菜种子发育过程中主要脂肪酸的积累及相关分析[J].江苏农业学报,2011,27(2):258-263.