摘要:为明确桑树在不同氮形态下光合能力差异以及桑树对不同氮形态的需求规律,以1年生桑树实生苗为试验材料,研究了在硝态氮![]() 铵态氮
铵态氮![]() 以及硝酸铵(NH4NO3)(氮素浓度均为7.5 mmol/L)处理下桑树叶片的叶绿素荧光特性。结果表明,桑树叶片在
以及硝酸铵(NH4NO3)(氮素浓度均为7.5 mmol/L)处理下桑树叶片的叶绿素荧光特性。结果表明,桑树叶片在![]() 下的PS Ⅱ光化学活性、电子传递速率和光能利用能力明显低于
下的PS Ⅱ光化学活性、电子传递速率和光能利用能力明显低于![]() 和NH4NO3处理,而
和NH4NO3处理,而![]() 和NH4NO3处理之间无明显差异
和NH4NO3处理之间无明显差异![]() 处理下桑树叶片的VJ和VI均较
处理下桑树叶片的VJ和VI均较![]() 和NH4NO3处理明显增加,即PS Ⅱ反应中心受体侧电子QA向QB传递速率较低,但此时桑树叶片的Sm和N却明显高于
和NH4NO3处理明显增加,即PS Ⅱ反应中心受体侧电子QA向QB传递速率较低,但此时桑树叶片的Sm和N却明显高于![]() 和NH4NO3处理,说明导致
和NH4NO3处理,说明导致![]() 处理下桑树叶片PS Ⅱ受体侧电子传递能力降低的原因直接与QB功能的降低有关。另外
处理下桑树叶片PS Ⅱ受体侧电子传递能力降低的原因直接与QB功能的降低有关。另外![]() 处理下桑树叶片的VK和VL也明显低于
处理下桑树叶片的VK和VL也明显低于![]() 和NH4NO3处理,说明桑树叶片OEC活性的抑制与类囊体膜结构的稳定性降低也是导致其叶片PS Ⅱ反应中心光化学活性在
和NH4NO3处理,说明桑树叶片OEC活性的抑制与类囊体膜结构的稳定性降低也是导致其叶片PS Ⅱ反应中心光化学活性在![]() 处理下降低的重要原因。
处理下降低的重要原因。
关键词:桑树;氮形态;叶绿素荧光;PSⅡ
 ).The results showed that the photochemical activity,electron transfer rate and light energy utilization of mulberry leaves in PSⅡ
).The results showed that the photochemical activity,electron transfer rate and light energy utilization of mulberry leaves in PSⅡ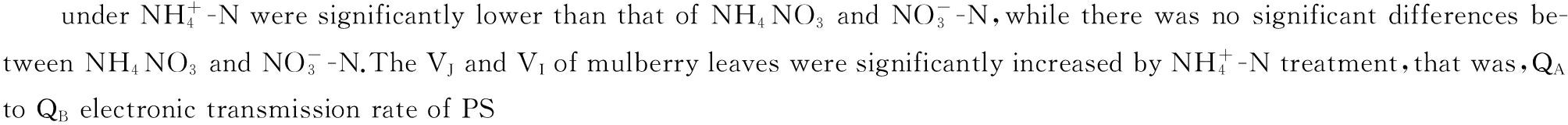 Ⅱ
Ⅱ![]() Ⅱ
Ⅱ Ⅱ
Ⅱ![]() t.
t.
氮是植物体最为重要的元素之一,植物体内一半以上的氮主要以酶的形式存在于植物的叶绿体内,参与植物的光合代谢过程[1],在植物的生长发育中起到重要的作用[2]。植物除了少量吸收氨基酸外,根系对氮的吸收主要以硝态氮![]() 铵态氮
铵态氮![]() 和有机态氮为主[3-4],并且
和有机态氮为主[3-4],并且![]() 或
或![]() 是主要的吸收形式[5]。不同植物种类对氮的偏好也不相同,如白云杉(Picea glauca)[6]、欧洲赤松(Pinus sylvestris)[7]和水稻(Oryza sativa L.)[8]相对喜欢
是主要的吸收形式[5]。不同植物种类对氮的偏好也不相同,如白云杉(Picea glauca)[6]、欧洲赤松(Pinus sylvestris)[7]和水稻(Oryza sativa L.)[8]相对喜欢![]() 而红松(Pinus koraiensis Sieb.et Zucc.)[9]和烟草(Nicotiana tabacum L.)[10]等喜
而红松(Pinus koraiensis Sieb.et Zucc.)[9]和烟草(Nicotiana tabacum L.)[10]等喜![]() 植物在
植物在![]() 存在过多时,会发生毒害现象[11]。与
存在过多时,会发生毒害现象[11]。与![]() 相比,相对增加
相比,相对增加![]() 可明显提高芥菜(Indian mustard)[12]和油菜(Brassica campestris L.)[13]的抗逆性,而盐胁迫下对大麦(Hordeum L.)施用
可明显提高芥菜(Indian mustard)[12]和油菜(Brassica campestris L.)[13]的抗逆性,而盐胁迫下对大麦(Hordeum L.)施用![]() 则更为有利[14]。
则更为有利[14]。
桑树因具有较强的抗逆能力,不但具有重要的经济价值,还可以作为退化生态地区植被恢复的优良树种[15-16],但在退化生态地区土壤中氮缺乏是限制植物生长,导致其植被恢复的困难因素之一[17-18],合理施用氮肥是提高退化地植被恢复的重要措施之一。桑树为喜![]() 植物,在
植物,在![]() 的供应下,桑树叶片的光合能力和电子传递速率明显高于供应
的供应下,桑树叶片的光合能力和电子传递速率明显高于供应![]() 并且相对增施
并且相对增施![]() 还可以提高桑树的抗盐性[20]。但是关于不同氮形态下桑树叶片PSⅡ光化学活性,特别是对桑树在
还可以提高桑树的抗盐性[20]。但是关于不同氮形态下桑树叶片PSⅡ光化学活性,特别是对桑树在![]() 作用下具有较高电子传递速率及光合能力的原因尚不明确。为此,本研究利用叶绿素荧光技术以及快相叶绿素荧光动力学(OJIP曲线)方法研究了桑树幼苗叶片PSⅡ光化学活性、电子供体侧放氧复合体OEC的功能、电子受体侧电子传递能力、电子库容量、类囊体膜状态以及光能分配参数等对不同氮素形态的生理响应,探讨了桑树在
作用下具有较高电子传递速率及光合能力的原因尚不明确。为此,本研究利用叶绿素荧光技术以及快相叶绿素荧光动力学(OJIP曲线)方法研究了桑树幼苗叶片PSⅡ光化学活性、电子供体侧放氧复合体OEC的功能、电子受体侧电子传递能力、电子库容量、类囊体膜状态以及光能分配参数等对不同氮素形态的生理响应,探讨了桑树在![]() 供应下光合能力大于
供应下光合能力大于![]() 的原因,对深入研究桑树的合理施肥提供一些基础数据。
的原因,对深入研究桑树的合理施肥提供一些基础数据。
1.1 试验材料及处理方法
试验于2015年3-6月在东北农业大学进行。供试桑树材料为东北农业大学土壤学实验室自育的当年播种实生苗。待幼苗长至约30 cm时,进行水培养,水培养营养液为1/2Hoagland完全营养液。水培箱规格:直径30 cm,高50 cm,不透光。培养条件:温度(25±3)℃、光照强度400 μmol/(m2·s)、光周期12 h/12 h(光/暗),水培时电动气泵连续通气,每3 d更换一次营养液。水培30 d幼苗缓苗后,进行不同形态氮素试验处理。
试验共设单一![]() 浓、单一
浓、单一![]() 和NH4NO3 3个处理,不同处理氮的总浓度均设为
和NH4NO3 3个处理,不同处理氮的总浓度均设为![]() 处理即为培养时的1/2Hoagland完全营养液
处理即为培养时的1/2Hoagland完全营养液![]() 处理和NH4NO3处理分别将Hoagland完全营养液中的Ca(NO3)2·4H2O分别以同等氮浓度的(NH4)2SO4和NH4NO3进行替换
处理和NH4NO3处理分别将Hoagland完全营养液中的Ca(NO3)2·4H2O分别以同等氮浓度的(NH4)2SO4和NH4NO3进行替换![]() 和NH4NO3处理中替换的Ca(NO3)2·4H2O中Ca2+浓度以CaCl2补充到
和NH4NO3处理中替换的Ca(NO3)2·4H2O中Ca2+浓度以CaCl2补充到![]() 处理中的Ca2+浓度水平。待不同氮形态处理30 d后进行各生理指标的测定。
处理中的Ca2+浓度水平。待不同氮形态处理30 d后进行各生理指标的测定。
1.2 测定参数和方法
叶绿素荧光参数及其光响应曲线的测定:利用暗适应夹对桑树幼苗顶端第2片完全展开叶片进行0.5 h的暗适应,采用便携式脉冲调制荧光仪FMS-2(Hansatch公司,英国)测定暗适应后PSⅡ最大光化学效率(Fv/Fm),然后测定光下叶绿素荧光参数:实际光化学效率(ФPSⅡ)、光化学淬灭系数(qp)和非光化学淬灭(NPQ)等,每次测定重复3次。利用FMS-2的内置光源分别测定经0.5 h暗适应后桑树幼苗叶片的ФPSⅡ和ETR光响应曲线,FMS-2施加作用光强分别为100,200,400,600,800,1 000,1 200 μmol/(m2·s),每个作用光强下适应3 min[21]。
快相叶绿素荧光动力学曲线的测定:选择顶端第2片完全展开叶片,进行30 min暗适应后利用Mini调制式掌上叶绿素荧光仪(FluorPen FP 100 max,捷克)测定各暗适应后叶片的OJIP曲线。OJIP曲线由3 000 μmol/(m2·s)的脉冲红光诱导,相对荧光强度的记录从10 μs开始至1 s结束。其中OJIP曲线上O、J、I和P点分别为0,2,30和1 000 ms对应的时刻,L和K点分别表示0.15,0.30 ms对应的时刻。为分析J、I、L和K点相对可变荧光VJ、VI、VL和VK的变化,分别将O-P、O-J和O-K曲线标准化[22]。按照Strasser等[23]的方法对OJIP曲线进行JIP-test分析,得到QA完全被还原所需要的能量(Sm)、QA被氧化还原次数(N)、吸收光能用于QA-以后的电子传递的能量比例(φEo)和非光化学淬灭的最大量子产额(φDo)等参数。OJIP曲线的测定每个处理重复5次。
1.3 数据处理方法
运用Excel和SPSS(22.0)软件对测定数据进行统计分析,并采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)比较不同数据组间的差异。
2.1 桑树幼苗叶片叶绿素荧光参数对不同氮素形态的响应
由图1可以看出![]() 处理下桑树叶片的Fv/Fm、ФPS Ⅱ和qp分别低于
处理下桑树叶片的Fv/Fm、ФPS Ⅱ和qp分别低于![]() 处理9.13%(P<0.01)、55.53%(P<0.01)和26.27%(P<0.01),分别低于NH4NO3处理10.65%(P<0.01),55.36%(P<0.01)和38.87%(P<0.01),均达极显著差异水平。而
处理9.13%(P<0.01)、55.53%(P<0.01)和26.27%(P<0.01),分别低于NH4NO3处理10.65%(P<0.01),55.36%(P<0.01)和38.87%(P<0.01),均达极显著差异水平。而![]() 处理下桑树叶片的NPQ则明显高于
处理下桑树叶片的NPQ则明显高于![]() 和NH4NO3处理
和NH4NO3处理![]() 和NH4NO3处理下桑树叶片的各叶绿素荧光参数之间均无显著差异。
和NH4NO3处理下桑树叶片的各叶绿素荧光参数之间均无显著差异。
图中数据为平均值±标准差;不同小写字母表示差异显著,不同大写字母表示差异极显著。图4-5同。
Data in the figure are mean±SE:Values followed by different small letters mean significant difference(P<0.05);
Values followed by different capital letters mean significant difference(P<0.01).The same as Fig.4-5.
图1 桑树幼苗叶片叶绿素荧光特性对不同氮素形态的响应
Fig.1 Responses of chlorophyll fluorescence characteristics in leaves of Morus alba seedlings to nitrogen form
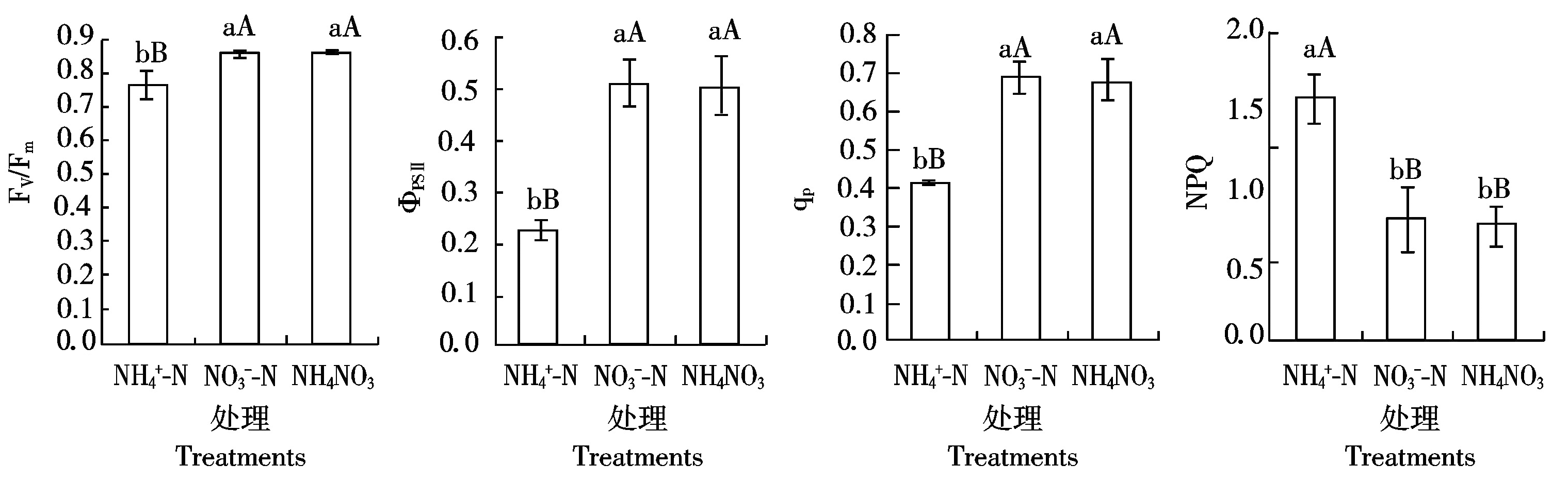
2.2 不同氮形态下桑树幼苗叶片叶绿素荧光参数的光响应曲线
由图2可以看出,随着光强的增加,桑树幼苗叶片的ФPSⅡ呈逐渐降低趋势,而ETR则呈先增加后降低趋势,存在明显的光饱和现象。但是不同处理之间明显不同,在不同光强下![]() 和NH4NO3处理桑树叶片的ФPSⅡ和ETR均无明显差异,但不同光强下
和NH4NO3处理桑树叶片的ФPSⅡ和ETR均无明显差异,但不同光强下![]() 处理桑树叶片的ФPSⅡ均明显低于
处理桑树叶片的ФPSⅡ均明显低于![]() 和NH4NO3处理,并且除100,200 μmol/(m2·s)光强下3个处理叶片ETR无明显差异外,当光强超过200 μmol/(m2·s)后
和NH4NO3处理,并且除100,200 μmol/(m2·s)光强下3个处理叶片ETR无明显差异外,当光强超过200 μmol/(m2·s)后![]() 处理下桑树叶片的ETR也明显低于
处理下桑树叶片的ETR也明显低于![]() 和NH4NO3处理。另外
和NH4NO3处理。另外![]() 和NH4NO3处理下桑树叶片的ETR在光强超过1 000 μmol/(m2·s)后呈降低趋势,而
和NH4NO3处理下桑树叶片的ETR在光强超过1 000 μmol/(m2·s)后呈降低趋势,而![]() 处理下则在光强为800 μmol/(m2·s)左右时就达到电子传递速率的光饱和点。
处理下则在光强为800 μmol/(m2·s)左右时就达到电子传递速率的光饱和点。
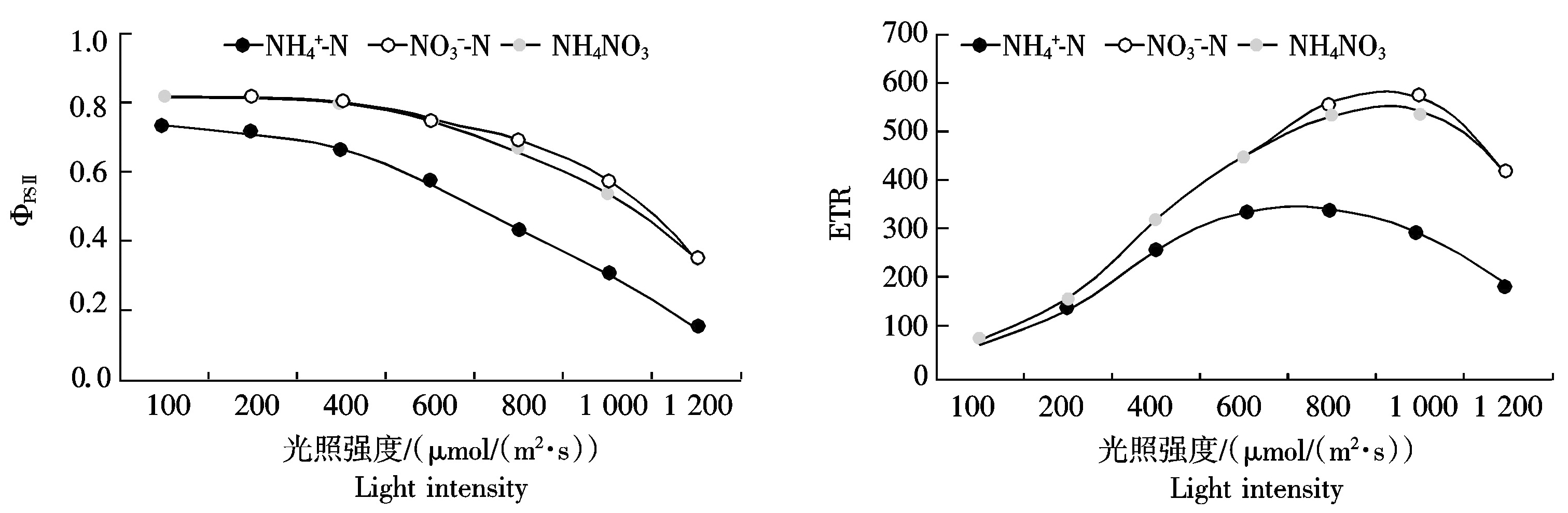
图2 不同氮形态下桑树幼苗叶片叶绿素荧光参数的光响应曲线
Fig.2 Chlorophyll fluorescence parameters response to illumination intensity in leaves of Morus alba seedlings under different nitrogen form
2.3 桑树幼苗叶片OJIP曲线对不同氮素形态的响应
由图3-A可以看出,不同氮形态处理下桑树叶片的OJIP曲线明显不同![]() 和NH4NO3处理下,OJIP曲线上O到J点的相对荧光强度无明显差异,而随着时间的延长,到达I点和P点时,NH4NO3处理下桑树叶片的相对荧光强度逐渐低于
和NH4NO3处理下,OJIP曲线上O到J点的相对荧光强度无明显差异,而随着时间的延长,到达I点和P点时,NH4NO3处理下桑树叶片的相对荧光强度逐渐低于![]() 处理
处理![]() 处理下桑树叶片O点和J点的相对荧光强度明显高于
处理下桑树叶片O点和J点的相对荧光强度明显高于![]() 和NH4NO3处理,但P点的相对荧光强度则分别低于
和NH4NO3处理,但P点的相对荧光强度则分别低于![]() 和NH4NO3处理。
和NH4NO3处理。
分别将OJIP曲线按O-P、O-J和O-K标准化后可以看出![]() 处理下桑树叶片J点(2 ms)和I点(30 ms)相对可变荧光均较
处理下桑树叶片J点(2 ms)和I点(30 ms)相对可变荧光均较![]() 和NH4NO3处理明显增加,并且NH4NO3处理下桑树叶片J点相对可变荧光稍大于
和NH4NO3处理明显增加,并且NH4NO3处理下桑树叶片J点相对可变荧光稍大于![]() 处理,而I点则无明显差异(图
处理,而I点则无明显差异(图![]() 处理下桑树叶片K点(0.3 ms)相对可变荧光明显高于
处理下桑树叶片K点(0.3 ms)相对可变荧光明显高于![]() 和NH4NO3处理,而
和NH4NO3处理,而![]() 和NH4NO3处理之间无明显差异(图3-C)。不同处理下标准化O-K曲线差异较小,仅表现为
和NH4NO3处理之间无明显差异(图3-C)。不同处理下标准化O-K曲线差异较小,仅表现为![]() 处理下桑树叶片L点(0.15 ms)相对可变荧光稍高于
处理下桑树叶片L点(0.15 ms)相对可变荧光稍高于![]() 和NH4NO3处理(图3-D)。
和NH4NO3处理(图3-D)。
2.4 桑树幼苗叶片OJIP曲线上各特征点相对可变荧光对不同氮素形态的响应
定量分析各点相对可变荧光之间差异可以看出(图![]() 处理下桑树叶片L、K、J和I点的相对可变荧光VL、VK、VJ和VI均较
处理下桑树叶片L、K、J和I点的相对可变荧光VL、VK、VJ和VI均较![]() 和NH4NO3处理明显提高,分别高于
和NH4NO3处理明显提高,分别高于![]() 处理7.19%(P>0.05),29.97%(P<0.01),46.39%(P<0.01)和15.02%(P<0.01),分别高于NH4NO3处理10.38%(P>0.05),31.43%(P<0.01),31.38%(P<0.01)和19.83%(P<0.01),除VL差异不显著外,VK、VJ和VI均达极显著差异水平
处理7.19%(P>0.05),29.97%(P<0.01),46.39%(P<0.01)和15.02%(P<0.01),分别高于NH4NO3处理10.38%(P>0.05),31.43%(P<0.01),31.38%(P<0.01)和19.83%(P<0.01),除VL差异不显著外,VK、VJ和VI均达极显著差异水平![]() 和NH4NO3处理之间的相对可变荧光差异较小
和NH4NO3处理之间的相对可变荧光差异较小![]() 处理下VI较NH4NO3处理提高了4.13%(P<0.05),达显著差异水平。
处理下VI较NH4NO3处理提高了4.13%(P<0.05),达显著差异水平。
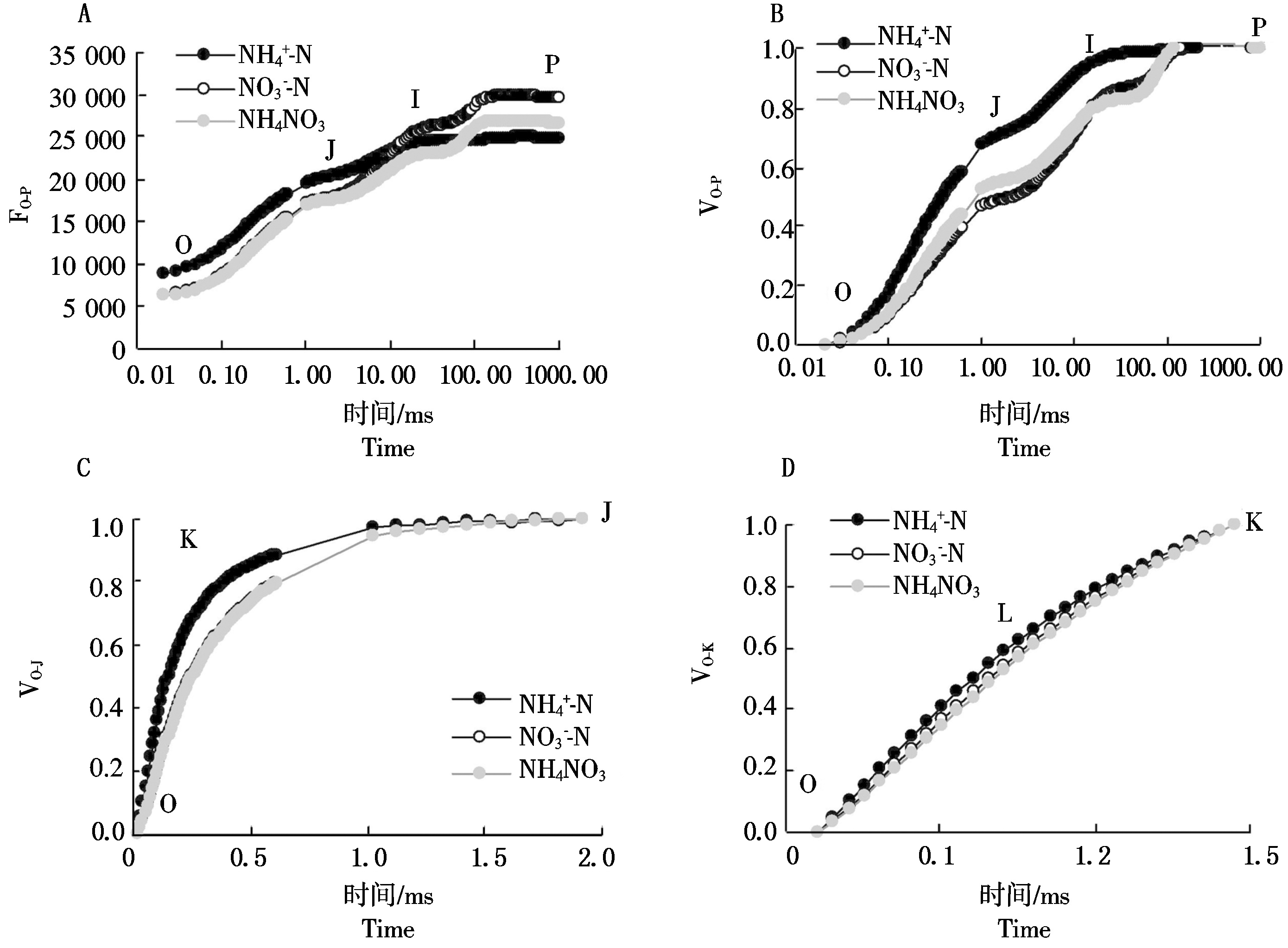
图3 桑树幼苗叶片OJIP曲线对不同氮素形态的响应
Fig.3 Responses of OJIP curve in leaves of Morus alba seedlings to nitrogen form
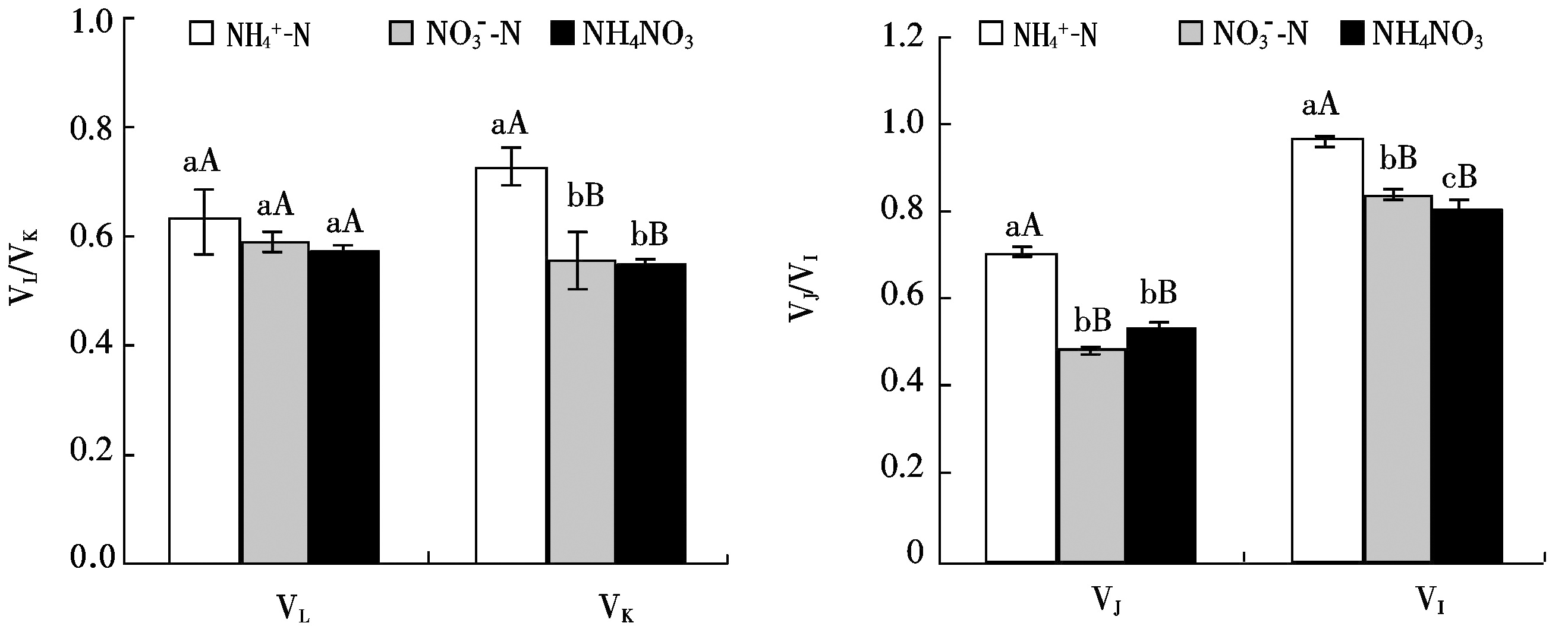
图4 桑树幼苗叶片VL、VK、VJ 和VI对不同氮素形态的响应
Fig.4 Responses of VL,VK,VJ and VI in leaves of Morus alba seedlings to nitrogen form
2.5 桑树幼苗叶片Sm、N、φEo和φDo对不同氮素形态的响应
由图5可以看出,不同氮形态下桑树叶片Sm和N表现出相似的变化规律,即![]() 处理下桑树叶片的Sm和N极显著高于
处理下桑树叶片的Sm和N极显著高于![]() 和NH4NO3处理,而NH4NO3处理较
和NH4NO3处理,而NH4NO3处理较![]() 处理稍有降低,但差异不显著
处理稍有降低,但差异不显著![]() 处理下桑树叶片的φEo分别低于
处理下桑树叶片的φEo分别低于![]() 和NH4NO3处理54.27%(P<0.01)和
和NH4NO3处理54.27%(P<0.01)和![]() 处理桑树叶片的φE0,高于NH4NO3处理
处理桑树叶片的φE0,高于NH4NO3处理![]() 处理下桑树叶片的φDo分别高于
处理下桑树叶片的φDo分别高于![]() 和NH4NO3处理68.64%(P<0.01)和
和NH4NO3处理68.64%(P<0.01)和![]() 和NH4NO3处理下桑树叶片的φDo无显著差异。
和NH4NO3处理下桑树叶片的φDo无显著差异。
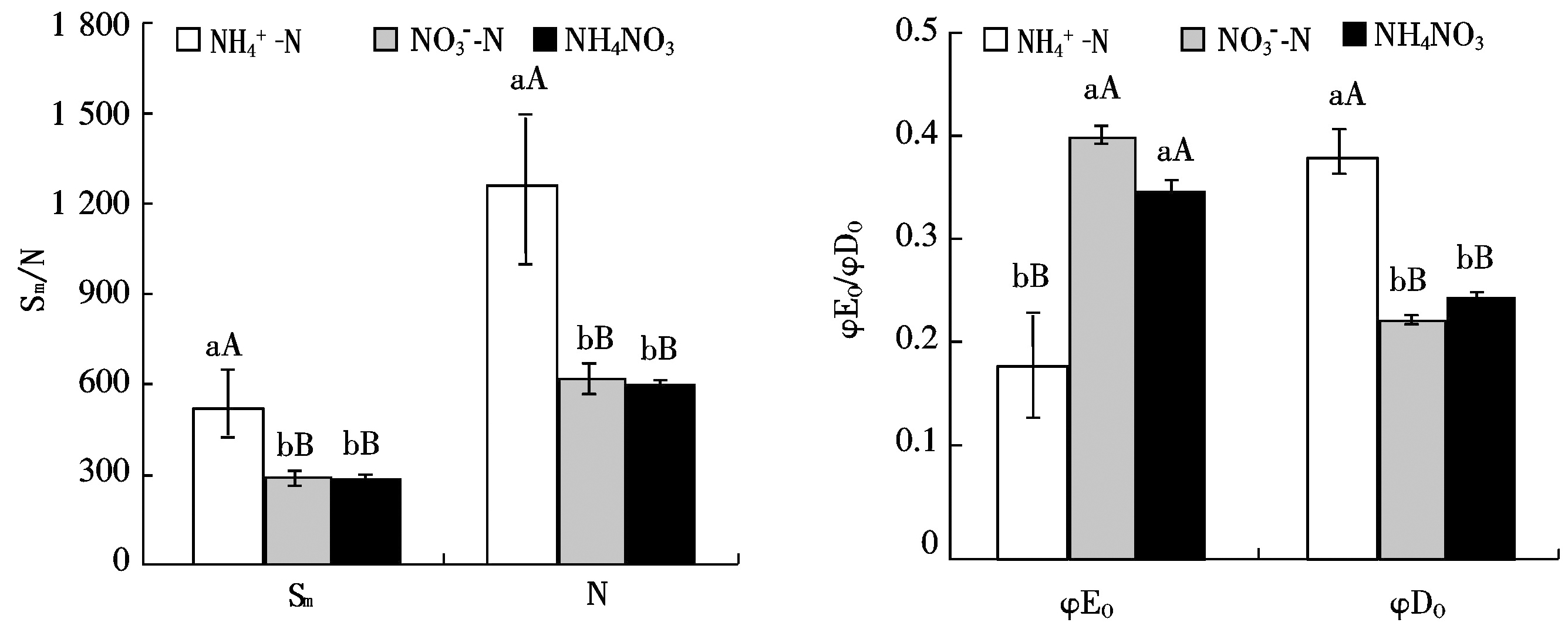
图5 桑树幼苗叶片Sm、N、φEo和φDo对不同氮素形态的响应
Fig.5 Responses of Sm,N,φEo and φDo in leaves of Morus alba seedlings to nitrogen form
叶绿素荧光技术在分析植物对光能的吸收和利用以及植物光系统功能方面发挥重要的作用。不同氮素形态对植物光合作用的影响明显不同,陈贵等[24-25]的研究表明,水分胁迫下![]() 供应对水稻光合效率的抑制效应小于
供应对水稻光合效率的抑制效应小于![]() 供应,叶片中Rubisco含量较高,而单施
供应,叶片中Rubisco含量较高,而单施![]() 则可以提高柚(Citrus maxima)在CO2倍增下的净光合速率[26]。本试验中
则可以提高柚(Citrus maxima)在CO2倍增下的净光合速率[26]。本试验中![]() 处理下桑树叶片的Fv/Fm、ФPS Ⅱ和qP均极显著低于
处理下桑树叶片的Fv/Fm、ФPS Ⅱ和qP均极显著低于![]() 处理,并且通过叶绿素荧光参数的光响应曲线也可以看出,不同光强下
处理,并且通过叶绿素荧光参数的光响应曲线也可以看出,不同光强下![]() 处理下桑树叶片的ФPSⅡ和ETR均明显低于
处理下桑树叶片的ФPSⅡ和ETR均明显低于![]() 处理,并且
处理,并且![]() 处理下桑树叶片的ETR光饱和点明显低于
处理下桑树叶片的ETR光饱和点明显低于![]() 处理。说明
处理。说明![]() 处理下桑树叶片的PSⅡ光化学活性和光能利用能力明显低于
处理下桑树叶片的PSⅡ光化学活性和光能利用能力明显低于![]() 处理,但是
处理,但是![]() 处理下桑树叶片的NPQ则明显高于
处理下桑树叶片的NPQ则明显高于![]() 处理,说明
处理,说明![]() 处理下桑树叶片主要通过启动依赖于叶黄素循环的非辐射能量耗散机制来降低其叶片中过剩激发能,但这同样也降低了光能用于光化学反应的比例,这会直接导致
处理下桑树叶片主要通过启动依赖于叶黄素循环的非辐射能量耗散机制来降低其叶片中过剩激发能,但这同样也降低了光能用于光化学反应的比例,这会直接导致![]() 处理下桑树叶片同化力(ATP和NADPH)的供应,进而抑制其光合碳同化能力导致同化物的积累量降低。这与众多的研究结果中单一的
处理下桑树叶片同化力(ATP和NADPH)的供应,进而抑制其光合碳同化能力导致同化物的积累量降低。这与众多的研究结果中单一的![]() 会对植物产生毒害现象一
会对植物产生毒害现象一![]() 和
和![]() 的合理配施可以明显缓解植物的铵盐毒害现象,Jan等[28]的研究发现
的合理配施可以明显缓解植物的铵盐毒害现象,Jan等[28]的研究发现![]() 与
与![]() 混合施用,与单纯
混合施用,与单纯![]() 相比,植物的干物质积累和蛋白质含量明显提高。Nathawat等[29]的研究发现
相比,植物的干物质积累和蛋白质含量明显提高。Nathawat等[29]的研究发现![]() 和
和![]() 混合施用与
混合施用与![]() 相比,印度芥菜(Brassica juncea L.)的种子产量无明显差异,但均明显高于
相比,印度芥菜(Brassica juncea L.)的种子产量无明显差异,但均明显高于![]() 处理;同时供应
处理;同时供应![]() 和
和![]() 也更有利于柑橘(Citrus reticulata)生长。本试验中,NH4NO3处理下桑树叶片的各叶绿素荧光参数均明显高于
也更有利于柑橘(Citrus reticulata)生长。本试验中,NH4NO3处理下桑树叶片的各叶绿素荧光参数均明显高于![]() 处理,并且与
处理,并且与![]() 基本无明显差异,NH4NO3处理即
基本无明显差异,NH4NO3处理即![]() 和
和![]() 处理明显缓解了桑树的铵盐毒害现象。
处理明显缓解了桑树的铵盐毒害现象。
快相叶绿素荧光动力学技术的应用,可以定性和定量分析光合作用中系统不同部位的功能,对深入研究植物的光合功能具有重要的意义。本试验结果中,不同氮形态处理下桑树叶片的OJIP曲线明显不同,其中![]() 处理下桑树叶片O点和J点的相对荧光强度明显高于
处理下桑树叶片O点和J点的相对荧光强度明显高于![]() 和NH4NO3处理,但P点的相对荧光强度则分别低于
和NH4NO3处理,但P点的相对荧光强度则分别低于![]() 和NH4NO3处理,即
和NH4NO3处理,即![]() 处理下桑树叶片的OJIP曲线更为平缓,这也是其PSⅡ反应中心活性较低的重要标志。但由于原始的OJIP曲线受环境影响较大,其相对荧光强度受多种环境因素共同作用,因此,常常将OJIP曲线进行标准化处理。本试验中,分别将OJIP曲线按O-P、O-J和O-K标准化后可以看出
处理下桑树叶片的OJIP曲线更为平缓,这也是其PSⅡ反应中心活性较低的重要标志。但由于原始的OJIP曲线受环境影响较大,其相对荧光强度受多种环境因素共同作用,因此,常常将OJIP曲线进行标准化处理。本试验中,分别将OJIP曲线按O-P、O-J和O-K标准化后可以看出![]() 处理下桑树叶片J点(2 ms)和I点(30 ms)相对可变荧光VJ和VI均较
处理下桑树叶片J点(2 ms)和I点(30 ms)相对可变荧光VJ和VI均较![]() 和NH4NO3处理明显增加,并且NH4NO3处理下桑树叶片J点相对可变荧光稍大于
和NH4NO3处理明显增加,并且NH4NO3处理下桑树叶片J点相对可变荧光稍大于![]() 处理,而I点则无明显差异。VJ反映了PSⅡ反应中心受体侧QA向QB传递情况,而I点相对可变荧光的增加则是因为QA-向QB传递过程中受到抑制导致[30]。因此,本试验结果说明
处理,而I点则无明显差异。VJ反映了PSⅡ反应中心受体侧QA向QB传递情况,而I点相对可变荧光的增加则是因为QA-向QB传递过程中受到抑制导致[30]。因此,本试验结果说明![]() 处理下桑树叶片的PSⅡ反应中心受体侧电子QA向QB传递能力明显低于
处理下桑树叶片的PSⅡ反应中心受体侧电子QA向QB传递能力明显低于![]() 和NH4NO3处理,但是
和NH4NO3处理,但是![]() 处理下桑树叶片的Sm和N却明显高于
处理下桑树叶片的Sm和N却明显高于![]() 和NH4NO3处理,即PSⅡ反应中心受体侧PQ库的较大,QA-被还原的次数较多,因此,这也说明造成
和NH4NO3处理,即PSⅡ反应中心受体侧PQ库的较大,QA-被还原的次数较多,因此,这也说明造成![]() 处理下桑树叶片QA向QB传递能力较低的原因主要与QB接受电子能力的降低有关,而不是受其电子传递下游PQ库接受电子能力的限制,桑树叶片中QB功能的相对不稳定性已经在前期的试验中得到了证明[31]。
处理下桑树叶片QA向QB传递能力较低的原因主要与QB接受电子能力的降低有关,而不是受其电子传递下游PQ库接受电子能力的限制,桑树叶片中QB功能的相对不稳定性已经在前期的试验中得到了证明[31]。
![]() 处理下桑树叶片K点(0.3 ms)相对可变荧光VK明显高于
处理下桑树叶片K点(0.3 ms)相对可变荧光VK明显高于![]() 和NH4NO3处理,而
和NH4NO3处理,而![]() 和NH4NO3处理之间无明显差异,说明
和NH4NO3处理之间无明显差异,说明![]() 和NH4NO3处理下桑树叶片的OEC活性明显高于
和NH4NO3处理下桑树叶片的OEC活性明显高于![]() 处理,较高的OEC活性保证了电子传递供体侧水裂解功能的正常,为光合电子传递链提供足够的电子供应。而
处理,较高的OEC活性保证了电子传递供体侧水裂解功能的正常,为光合电子传递链提供足够的电子供应。而![]() 处理下桑树叶片OEC活性的降低,则可能导致水的不完全裂解,从而产生H2O2等活性氧,从而破坏其光合系统的正常功能,而电子传递受体侧QB接受电子能力的降低原因则可能直接与其受到活性氧的攻击有关。有研究表明
处理下桑树叶片OEC活性的降低,则可能导致水的不完全裂解,从而产生H2O2等活性氧,从而破坏其光合系统的正常功能,而电子传递受体侧QB接受电子能力的降低原因则可能直接与其受到活性氧的攻击有关。有研究表明![]() 使得番茄(Lycopersicon esculentum)叶片细胞超微结构发生变化,叶绿体肿胀、片层模糊,甚至出现断裂等现象,嗜锇颗粒明显积累,而NH4NO3则可明显改变这种状况[32],嗜锇颗粒是类囊体膜的解离的重要标志[33]。本试验结果表明,不同处理下标准化O-K曲线差异较小,但
使得番茄(Lycopersicon esculentum)叶片细胞超微结构发生变化,叶绿体肿胀、片层模糊,甚至出现断裂等现象,嗜锇颗粒明显积累,而NH4NO3则可明显改变这种状况[32],嗜锇颗粒是类囊体膜的解离的重要标志[33]。本试验结果表明,不同处理下标准化O-K曲线差异较小,但![]() 处理下桑树叶片L点(0.15 ms)相对可变荧光VL也明显高于
处理下桑树叶片L点(0.15 ms)相对可变荧光VL也明显高于![]() 和NH4NO3处理,即单一的
和NH4NO3处理,即单一的![]() 处理下桑树叶片类囊体的膜结构的稳定性较
处理下桑树叶片类囊体的膜结构的稳定性较![]() 和NH4NO3处理低。但是不同处理下VL的变化幅度均明显小于VK、VJ和VI,即不同氮形态下桑树叶片的类囊体结构的稳定性相对于OEC活性的PSⅡ受体侧电子传递功能而言是最不易受损伤的环节。
和NH4NO3处理低。但是不同处理下VL的变化幅度均明显小于VK、VJ和VI,即不同氮形态下桑树叶片的类囊体结构的稳定性相对于OEC活性的PSⅡ受体侧电子传递功能而言是最不易受损伤的环节。
桑树在![]() 处理下的PSⅡ光化学活性明显高于
处理下的PSⅡ光化学活性明显高于![]() 处理,单一的
处理,单一的![]() 对桑树有毒害作用,不但桑树叶片的PSⅡ光化学活性和电子传递能力降低,而且放氧复合体功能受到抑制,叶片吸收的光能主要以无效热能的形式耗散,用于光化学反应的比例降低,甚至类囊体结构发生了改变。但是
对桑树有毒害作用,不但桑树叶片的PSⅡ光化学活性和电子传递能力降低,而且放氧复合体功能受到抑制,叶片吸收的光能主要以无效热能的形式耗散,用于光化学反应的比例降低,甚至类囊体结构发生了改变。但是![]() 和NH4NO3处理桑树叶片的光合能力均明显高于
和NH4NO3处理桑树叶片的光合能力均明显高于![]() 处理,并且二者处理之间无明显差异,即
处理,并且二者处理之间无明显差异,即![]() 可以明显地缓解单一
可以明显地缓解单一![]() 下桑树的铵毒害现象。
下桑树的铵毒害现象。
参考文献:
[1] Shangguan Z P, Shao M A, Dyckmans J . Effect of nitrogen nutrition and water deficit on net photosynthetic rate and chlorophyll fluorescence in winter wheat[J]. Journal of Plant Physiology, 2000, 156(1): 46-51.
[2] Wang J, Baerenklau K A. Crop response functions integrating water, nitrogen, and salinity[J]. Agricultural Water Management,2014,139(3):17-30.
[3] Kuelland K. Amino acid absorption by arctic plants:implications for plant nutrition and nitrogen cycling[J]. Ecology 1994, 75(8): 2373-2383.
[4] Chapin F S, Autumn K, Pugnaire F. Evolution of suites of traits in response to environmental stress[J]. The American Naturalist, 1993, 142(142): 78-92.
[5] 王 磊,隆小华,郝连香,等.氮素形态对盐胁迫下菊芋幼苗PSⅡ光化学效率及抗氧化特性的影响[J].草业学报,2012,21(1):133-140.
[6] Kim T, Mills H A, Wetzstein H Y. Studies on effects of nitrogen form on growth, development, and nutrient uptake in pecan[J]. Journal of Plant Nutrition ,2002,25(3):497-508.
[7] Nordin A, Uggla C, Näsholm T. Nitrogen forms in bark, wood and foliage of nitrogen-fertilized Pinus sylvestris[J]. Tree Physiology, 2001, 21(1):59-64.
[8] 宋 娜,郭世伟,沈其荣.不同氮素形态营养及水分胁迫对分蘖期水稻水分吸收及光合特性的影响[J].南京农业大学学报,2006,29(4):64-69.
[9] 陈永亮,刘明河,李修岭.不同形态氮素配比对红松幼苗光合特性的影响[J].南京林业大学学报:自然科学版,2005,29(3):77-80.
[10] 杜 蕊,张会慧,田 野,等.氮素形态对烟草叶片光合PSⅡ能量分配比例的影响[J].湖北农业科学, 2014,53(15):3520-3524.
[11] 邹 娜,李保海,强晓敏,等.高铵胁迫对拟南芥根系向重性的影响及机制初探[J].土壤,2013,45(1):166-171.
[12] Nathawat N S, Kuhad M S, Goswami C L, et al. Interactive effect of N source on salinity on growth indices and ion content of Indian mustard[J]. Journal of Plant Nutrition,2007,30(4):569-598.
[13] 杨 瑛,马 梅,郑青松,等.不同供氮形态下油菜幼苗对盐胁迫的响应[J].植物营养与肥料学报,2012,18(5):1220-1227.
[14] Ali A, Tucker T C, Thompson T L, et al. Effects of salinity and mixed ammonium and nitrate nutrition on the growth and nitrogen utilization of barley[J]. Journal of Agronomy and Crop Science,2001,186(4):223-228.
[15] 张会慧,张秀丽,胡彦波,等.中碱钠盐胁迫对桑树幼苗生长及光合特性的影响[J].南京林业大学学报:自然科学版,2013,37(1):217-222.
[16] 张会慧,张秀丽,李 鑫,等.NaCl和Na2CO3胁迫对桑树幼苗生长和光合特性的影响[J].应用生态学报,2012,23(3):625-631.
[17] 原俊凤,田长彦,冯 固,等.硝态氮对盐胁迫下囊果碱蓬幼苗根系生长和耐盐性的影响[J].植物营养与肥料学报,2009,15(4) :953- 959.
[18] 王常慧,邢雪荣,韩兴国.草地生态系统中土壤氮素矿化影响因素的研究进展[J].应用生态学报,2004,15(11):2184-2188.
[19] 许 楠,张会慧,朱文旭,等.氮素形态对饲料桑树幼苗生长和光合特性的影响[J].草业科学, 2012, 29(10): 1568-1572.
[20] 逄好胜,张会慧,敖 红,等.硝态氮对Na2CO3胁迫下桑树幼苗生长和光合特性的影响[J].草业科学,2014,31(8):1515-1522.
[21] 张会慧,张秀丽,胡彦波,等.高粱-苏丹草杂交种的生长特性和光合功能研究[J].草地学报,2012,20(5):881-887.
[22] Zhang H H, Zhong H X, Wang J F, et al. Adaptive changes in chlorophyll content and photosynthetic features to low light in Physocarpus amurensis Maxim and Physocarpus opulifolius "Diabolo" [J].2016,4(3):e2125. DOI 10.7717/peerj.2125.
[23] Strasser R J, Srivastava A, Govindjee. Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria[J]. Photochemistry and Photobiology,1995,61(1):32-42.
[24] 陈 贵,周 毅,郭世伟,等.水分胁迫条件下不同形态氮素营养对水稻叶片光合效率的调控机理研究[J].中国农业科学,2007,40(10):2162-2168.
[25] 陈 贵,周 毅,郭世伟,等.局部根系水分胁迫下不同形态氮素营养对苗期水稻的影响[J].中国水稻科学,2006,20(6):638-644.
[26] 孙谷畴,曾小平,赵 平,等.不同氮源下生长的柚树叶片光合参数对高浓度CO2驯化作用的比较[J].生态学报,2003,23(1):14-21.
[27] ![]() ]. Proceedings of the National Academy of Sciences of the United States of America,2001,98(7):4255-4258.
]. Proceedings of the National Academy of Sciences of the United States of America,2001,98(7):4255-4258.
[28] Jan A, Osman M B, Amanullah. Response of chickpea to nitrogen sources under salinity stress[J]. Journal of Plant Nutrition,2013,36(9):1373-1382.
[29] Nathawat N S, Kuhad M S, Goswami C L, et al. Interactive effect of N source on salinity on growth indices and ion content of Indian mustard[J]. Journal of Plant Nutrition, 2007,30(4):569-598.
[30] 李鹏民,高辉远, Strasser R J.快速叶绿素荧光诱导动力学分析在光合作用研究中的应用[J].植物生理与分子生物学学报, 2005,31(6): 559-566.
[31] 张会慧,张秀丽,许 楠,等.盐胁迫下桑树叶片D1蛋白周转和叶黄素循环对PSⅡ的影响[J].林业科学,2013,49(1):99-106.
[32] 刘 冉,石 峰,刘伟成,等.不同形态氮素对盐胁迫下番茄细胞超微结构与光合作用的影响[J].园艺学报,2015,42(3):471-479.
[33] 马英姿,张 慧,王志毅,等.高温胁迫对蛇足石杉质膜透性及叶绿体结构的影响[J].中草药,2013,44(18):2605-2610.
欢迎订阅2017年《山西农业科学》
《山西农业科学》是山西省农业科学院主办的大农业学术性期刊(中国科技核心期刊),主要栏目有:宏观农业、生物技术、遗传育种、耕作栽培、生理生化、资源与环境、植物保护、畜牧兽医、水产渔业、贮藏与加工、信息技术、文献综述等。主要读者对象为:农业研究机构科研人员、农业院校师生、涉农部门农业技术推广工作者。
本刊为月刊,大16开本,96页码。每期定价8.00元,全年96.00元。国际刊号为ISSN 1002-2481,国内统一刊号CN 14-1113/S,邮发代号22-24。
欢迎订阅,欢迎投稿!
Key words:Mulberry;Nitrogen form;Chlorophyll fluorescence characteristics;PSⅡ
地址:太原市龙城大街81号 邮编:030031电话:0351-7089783E-mail:sxnykx@126.com
收稿日期:2017-01-07
基金项目:黑龙江省自然科学基金项目(C200917)
中图分类号:Q945.79;S143.1
文献标识码:A
文章编号:1000-7091(2017)02-0164-07
doi:10.7668/hbnxb.2017.02.025