粒重基因TaCwi-A1等位变异在黄淮麦区小麦品种(系)中的分布及功能分析
刘永伟1,周 硕1,王雪征2,孙果忠1,朱金永1,韩秋芬1,李春杰3,赵 和1,王海波1
(1.河北省农林科学院 遗传生理研究所,河北省植物转基因中心,河北 石家庄 050051;2.河北省农林科学院 旱作农业研究所,河北 衡水 053000;3.河北省种子管理总站,河北 石家庄 050031)
摘要:为探讨小麦粒重基因TaCwi-A1等位变异TaCwi-A1a和TaCwi-A1b在育种实践中的应用价值,首先利用TaCwi-A1功能标记对539份黄淮麦区小麦品种(系)进行分子检测,确定各供试材料的等位变异类型,从而获得TaCwi-A1a和TaCwi-A1b 2种等位变异的分布频率。对审定品种、参加区试品系以及自育品系进行了千粒质量(3个不同生长环境)及粒长、粒宽和籽粒面积(1个生长环境)测试分析,比较TaCwi-A1a和TaCwi-A1b等位变异籽粒表型性状的差异性。结果表明,在539份黄淮麦区小麦资源中TaCwi-A1a的分布频率为65.03%,TaCwi-A1b的分布频率为34.97%,TaCwi-A1a的分布频率明显高于TaCwi-A1b。籽粒表型性状分析表明,无论是审定品种,还是参加区试品系和自育品系,在3个生长环境下,TaCwi-A1a基因型材料的千粒质量均值都显著高于TaCwi-A1b;TaCwi-A1a基因型材料的粒长和粒宽均显著高于TaCwi-A1b。进一步验证了TaCwi-A1a等位变异的籽粒表型性状增效功能,说明其对粒重及其构成要素是优异的等位变异。此外,本研究鉴定了黄淮麦区中TaCwi-A1等位变异的分布情况,为亲本选配提供了参考。
关键词:普通小麦;TaCwi-A1;分子标记;等位变异;千粒质量;粒长;粒宽
小麦(Triticum aestivum L.)总产量的持续提高是保障我国粮食安全的战略需求。目前我国小麦种植面积逐年下降,要维持总产目标就需要不断提高单产水平。黄淮麦区是我国冬小麦主产区,常年小麦种植面积占全国小麦种植面积的40%~45%,产量占50%左右,品种在产量上的贡献占35%左右[1]。小麦高产取决于亩穗数、穗粒数和千粒质量三要素的突破与协调。已有研究表明,穗粒数多少受群体影响较大,而千粒质量则相对较独立[2]。近年来,小麦单产水平的不断提高与千粒质量呈现的逐渐递增有关[3]。但是,粒重是受多个微效基因控制的数量性状,仅靠表型选择聚合粒重优异等位基因难度会很大,分子标记辅助选择为我们提供了另一种育种方案。利用分子标记分析黄淮麦区小麦品种的遗传多样性,发掘种质资源中的粒重优异等位变异对于指导育种实践具有重要意义。
目前为止,已经克隆了一些小麦粒重相关基因并开发了功能标记。包括TaCwi-A1、TaSus2-2B、TaGW2、TaCKX6-D1、TaSAP1-A1、TaGS1a、TaGS-D1、和TaGASR7-A1等[4-13]。这些标记的开发对于了解小麦粒重调控机制和产量潜力的形成提供了有用的信息。其中,Ma等[4]利用水稻糖代谢相关的细胞壁转化酶基因CWI信息,克隆了普通小麦2A染色体上细胞壁转化酶基因TaCwi-A1的全长编码序列。并针对TaCwi-A1位点的等位变异TaCwi-Ala和TaCwi-Alb开发了显性互补标记CWI22和CWI21。通过对309份中国冬小麦推广品种和178份中国农家品种的检测,发现CWI21所扩增的404 bp条带与低千粒质量关联,而CWI22扩增的402 bp条带与高千粒质量关联。韩利明等[14]对来自21个国家的745份小麦品种资源进行了检测,发现TaCwi-A1基因的功能标记CWI22和CWI21能很好地区分等位变异TaCwi-A1a和TaCwi-A1b。相吉山等[15]用CWI22和CWI21标记检测了1241份新疆小麦品种资源,探讨了TaCwi-A1等位变异类型及分布频率,并利用其中110份冬小麦品种资源验证了CWI22和CWI21检测千粒质量的可靠性。上述研究都未将TaCwi-A1的等位变异与粒重构成要素进行详细的比较分析。
本研究以539份黄淮麦区的审定品种和参加各级别区试的品系等为研究对象,利用粒重基因TaCwi-A1功能标记CWI22和CWI21检测全部供试材料,获得其等位变异类型及分布频率,并对TaCwi-A1不同基因型材料的粒重及其构成要素进行比较分析。探讨等位变异TaCwi-A1a和TaCwi-A1b的实际育种价值,为小麦育种的亲本选配和分子标记辅助选择提供参考。
1 材料和方法
1.1 供试材料
材料为来源于我国黄淮冬麦区的河北、河南、山东、山西等省份的539份小麦品种资源。其中,包含审定品种107份,自育品系111份(其中有45份参加区试材料),其他为参加各级区域试验品种(系)321份。
供试材料于2013年10月至2015年6月连续2个生长季种植于河北省农林科学院鹿泉大河试验站,2014年10月至2015年6月种植于河北省农林科学院植物转基因中心试验地。记录环境1(E1):大河,2014;环境2(E2):大河,2015;环境3(E3):转基因中心,2015。试验材料顺序排列,双行区人工点播,行长2 m,行距25 cm,株距3 cm。田间管理方法同常规标准实验田。
1.2 小麦籽粒性状的测定
供试小麦材料成熟后及时收获晾干并脱粒。每个材料随机取500粒种子称重,每个材料2次重复,换算为该材料的千粒质量。粒长、粒宽和籽粒面积的测量参考张祖建等[16]图像扫描法得出。每个材料随机取种子200粒,经数粒板置于扫描仪面板上,使其有明显间隔,加标尺作为测距标准,形成图像文件。利用生物图像分析软件(Aon-Studio 2010)细胞分析模块测定各性状数据。
1.3 分子标记检测
采用CTAB法提取供试材料基因组DNA[17],每份材料提取3份,并用分光光度计检测浓度,以3份DNA的检测结果确定材料等位变异基因型。根据Ma等[4]开发的TaCwi-A1基因的一对显性互补标记CWI22和CWI21合成引物。引物序列如下:CWI22-F:5′-GGTGATGAGTTCATGGTTAAT-3′,CWI22-R:5′-AGAAGCCCAACATTAAATCAAC-3′。CWI21-F:5′-GTGGTGATGAGTTCATGGTTAAG-3′,CWI21-R:5′-AGAAGCCCAACATTAAATCAAC-3′。引物由上海捷瑞生物工程有限公司合成。PCR扩增体系为20 μL,包括1×PCR Buffer,200 μmol/L dNTPs,每条引物 10 pmol/L、1 unit Taq DNA polymerase以及40~60 ng模板DNA。反应程序:95 ℃预变性3 min;95 ℃变性30 s,60 ℃退火30 s,72 ℃延伸45 s,35个循环;72 ℃ 延伸10 min。在Eppendorf公司的Mastercycler Gradient梯度PCR仪进行扩增,扩增产物用2%的琼脂糖凝胶电泳和8%的聚丙烯酰胺凝胶电泳进行检测,凝胶成像拍照,统计扩增结果。
1.4 数据统计分析
数据统计分析使用Excel和SPSS 19.0(SPSS,Chicago,USA)软件。千粒质量、粒长、粒宽和籽粒面积等试验数据使用SPSS进行独立样本t检验(Independent-samples t test)。
2 结果与分析
2.1 黄淮麦区小麦品种资源中TaCwi-A1等位变异类型及分布频率
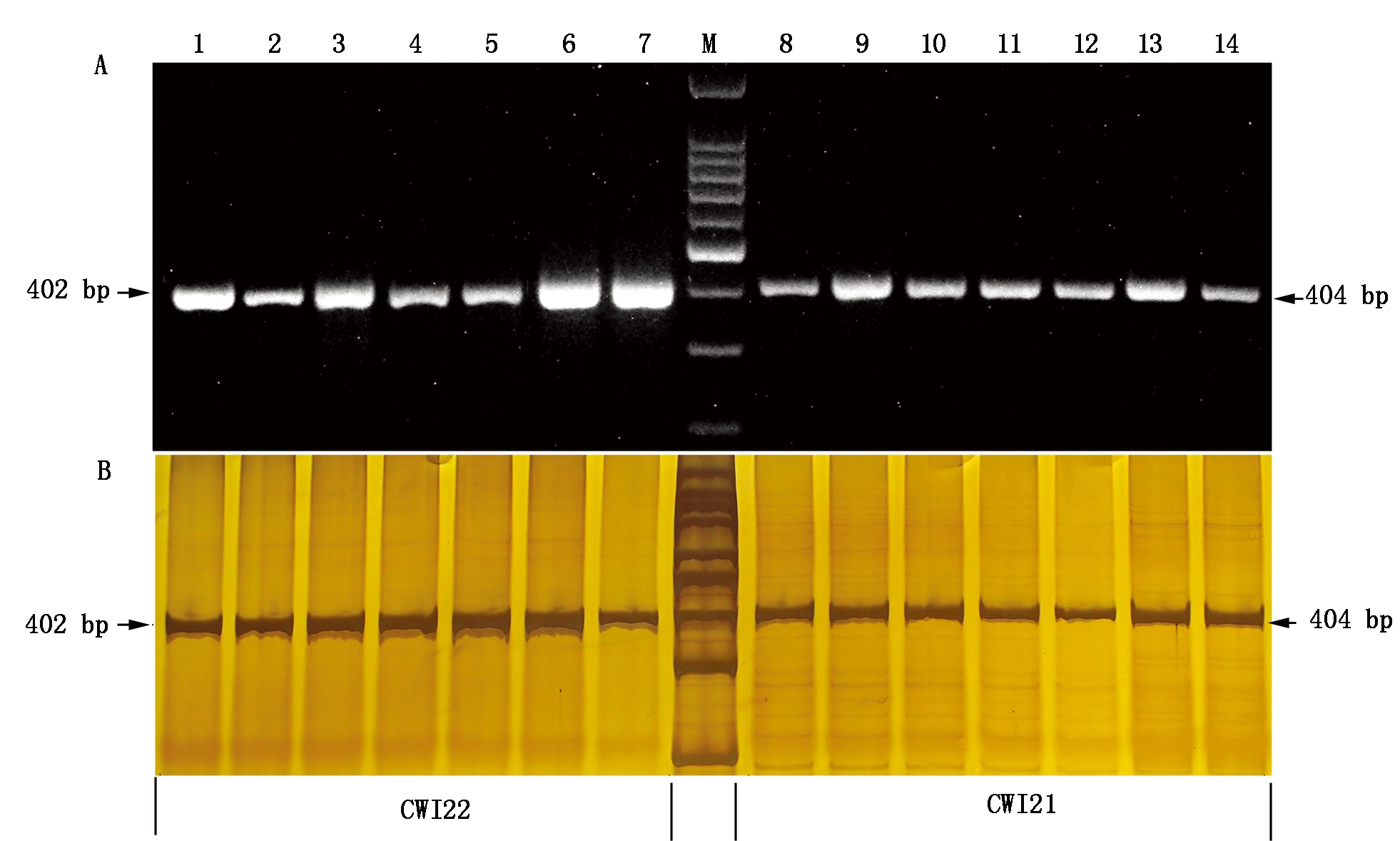
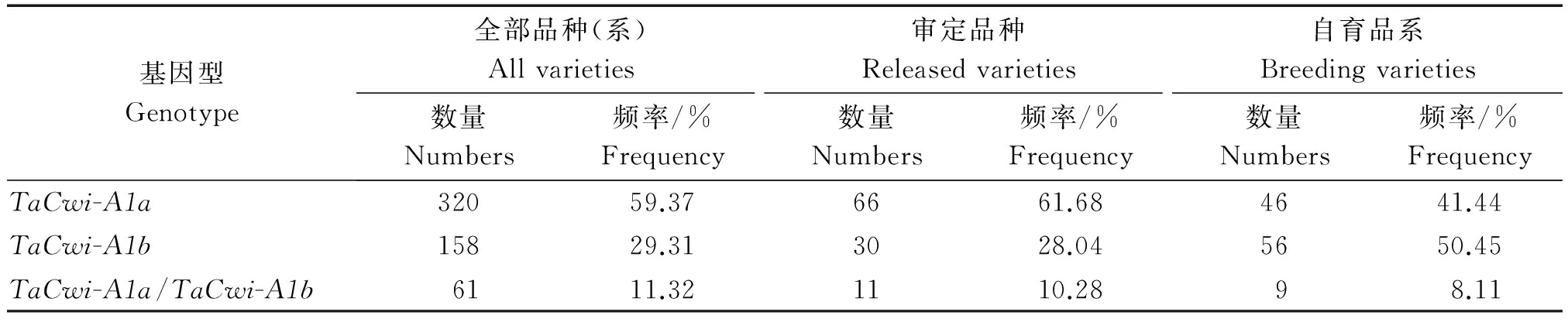
为了分析黄淮麦区小麦品种资源中TaCwi-A1等位变异类型及分布频率,利用小麦粒重基因TaCwi-A1的一对显性互补功能标记CWI22、CWI21对539份供试材料进行分子检测,结果表明(表1),539份黄淮麦区小麦品种资源中,有381份材料用CWI22能扩增出402 bp的目的片段,说明含有TaCwi-A1a;有219份材料能够用CWI21扩增出404 bp的目的片段,说明含有TaCwi-A1b(图1),其中61份材料用CWI22和CWI21都能扩增出特异条带,归为杂合类型。总体来看,TaCwi-A1a等位变异的分布频率为65.03%,TaCwi-A1b等位变异的分布频率为34.97%,TaCwi-A1a的分布频率明显高于TaCwi-A1b。
2.2 审定小麦品种中TaCwi-A1等位变异分布与籽粒表型性状分析
对107份黄淮麦区审定小麦品种中TaCwi-A1a、TaCwi-A1b等位变异进行了分子标记检测。结果表明,在审定品种中,TaCwi-A1a的分布频率为66.82%,TaCwi-A1b的分布频率为33.18%(表1)。
表1 黄淮小麦品种资源中TaCwi-A1a和TaCwi-A1b的分布频率
Tab.1 The frequencies of TaCwi-A1a and TaCwi-A1b in Yellow and Huai River Valley Winter Wheat Region varieties
基因型Genotype全部品种(系)Allvarieties审定品种Releasedvarieties自育品系Breedingvarieties数量Numbers频率/%Frequency数量Numbers频率/%Frequency数量Numbers频率/%FrequencyTaCwi-A1a32059.376661.684641.44TaCwi-A1b15829.313028.045650.45TaCwi-A1a/TaCwi-A1b6111.321110.2898.11
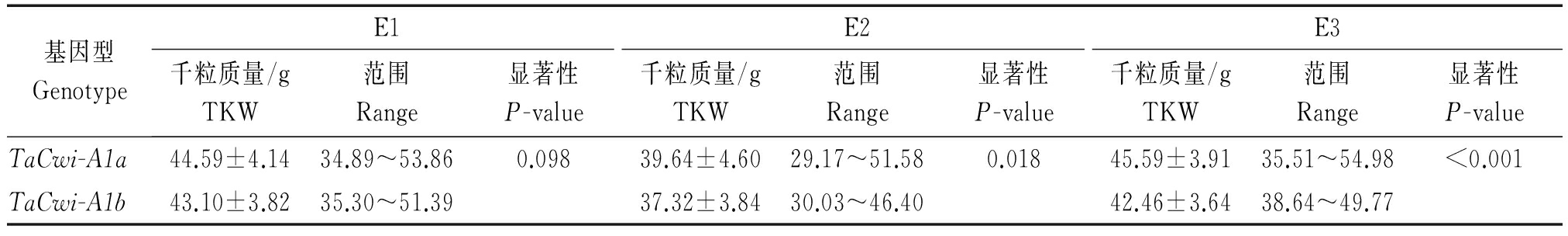
本研究还对审定品种中2种等位变异类型在不同环境条件下的千粒质量进行了分析(表2),t检验表明,在3个生长环境下,TaCwi-A1a基因型材料的千粒质量均高于TaCwi-A1b(E1的P值<0.10,E2的P值<0.05,E3的P值<0.001),平均千粒质量分别高1.49,2.32,3.13 g。这与Ma等[4]对2组中国冬小麦主栽品种和2组中国农家品种检测发现的标记与高低粒重的对应关系是一致的。说明CWI22、CWI21标记实用性较高,可用于小麦粒重性状的辅助选择。
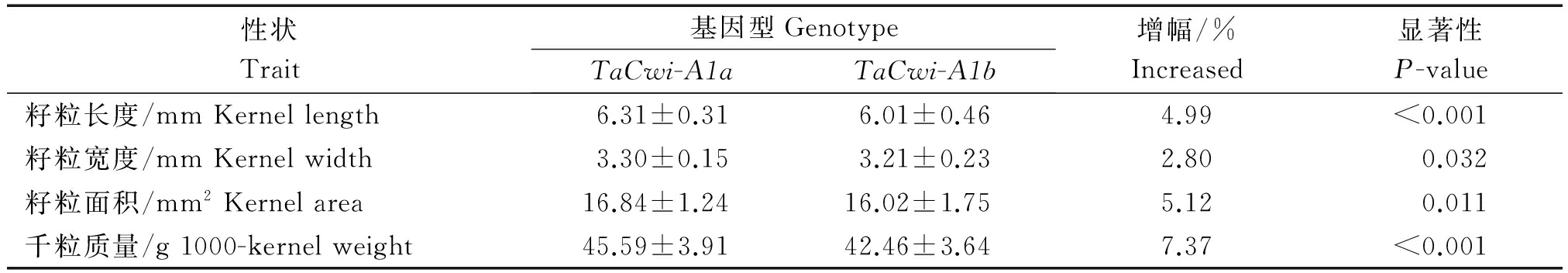
本研究还以E3环境下的表型数据为基础,对审定小麦品种中TaCwi-A1 2种等位变异与粒重各构成要素进行了关联分析(表3),结果表明,TaCwi-A1a基因型材料与TaCwi-A1b基因型材料的粒长(P<0.001)、粒宽(P<0.05)和籽粒面积(P<0.05)差异都达到显著水平。在粒重及其构成要素的增幅顺序是:千粒质量(7.37%)>籽粒面积(5.12%)>粒长(4.99%)>粒宽(2.80%)。可见,TaCwi-A1a基因型对应更高的籽粒表型性状数据,对粒重及其构成要素是优异的等位变异。
表2 审定品种中TaCwi-A1a和TaCwi-A1b在各环境下的千粒质量
Tab.2 The TKW of released varieties with TaCwi-A1a and TaCwi-A1b in three environments
基因型GenotypeE1E2E3千粒质量/gTKW范围Range显著性P-value千粒质量/gTKW范围Range显著性P-value千粒质量/gTKW范围Range显著性P-valueTaCwi-A1a44.59±4.1434.89~53.860.09839.64±4.6029.17~51.580.01845.59±3.9135.51~54.98<0.001TaCwi-A1b43.10±3.8235.30~51.3937.32±3.8430.03~46.4042.46±3.6438.64~49.77
表3 审定品种中TaCwi-A1等位变异的粒重表型性状比较
Tab.3 Comparison of grain phenotypic traits of TaCwi-A1 polymorphism in released varieties
性状Trait基因型GenotypeTaCwi-A1aTaCwi-A1b增幅/%Increased显著性P-value籽粒长度/mmKernellength6.31±0.316.01±0.464.99<0.001籽粒宽度/mmKernelwidth3.30±0.153.21±0.232.800.032籽粒面积/mm2Kernelarea16.84±1.2416.02±1.755.120.011千粒质量/g1000-kernelweight45.59±3.9142.46±3.647.37<0.001
2.3 参加区试的小麦品种资源中TaCwi-A1等位变异分布与千粒质量表型性状分析
对黄淮麦区河南、河北、山东3个主产区共计358份正在参加各级区域试验的材料中TaCwi-A1a和TaCwi-A1b等位变异进行了分子标记检测。结果表明,在河南省的品种资源中,TaCwi-A1a的分布频率为60.71%,TaCwi-A1b的分布频率为39.29%;在河北省的品种资源中,TaCwi-A1a的分布频率为65.00%,TaCwi-A1b的分布频率为35.00%;在山东省的品种资源中,TaCwi-A1a的分布频率为95.00%,TaCwi-A1b的分布频率为5.00%(表4)。这其中,由于河南、山东省的材料数量太少,统计数据仅供参考。但从整体趋势来看,黄淮麦区各育种单位选育的参试品系呈现TaCwi-A1a的分布频率明显高于TaCwi-A1b。
对上述材料中非杂合型的317份品系中2种等位变异类型在不同环境条件下的千粒质量表型值进行了分析(表5),t检验表明,TaCwi-A1a基因型材料的千粒质量显著高于TaCwi-A1b(E1的P值<0.10,E2的P值<0.05,E3的P值<0.05),平均千粒质量分别高0.93,1.06,1.02 g。
2.4 自育品系中TaCwi-A1等位变异分布与籽粒表型性状分析
通过上述对审定品种和参加区试品系的比较分析,我们可以看出TaCwi-A1基因的2种等位变异能够很好地区分高、低粒重性状。基于此,我们对111份自育品系中TaCwi-A1a、TaCwi-A1b等位变异进行了分子检测。结果表明,TaCwi-A1a的分布频率为45.50%,TaCwi-A1b的分布频率为54.50%(表1)。粒重优异等位变异TaCwi-A1a的分布频率低于河北省的平均分布频率65.00%。
表4 不同省份小麦品种资源中TaCwi-A1a和TaCwi-A1b的分布频率
Tab.4 The frequencies of TaCwi-A1a and TaCwi-A1b in three different provinces of wheat varieties
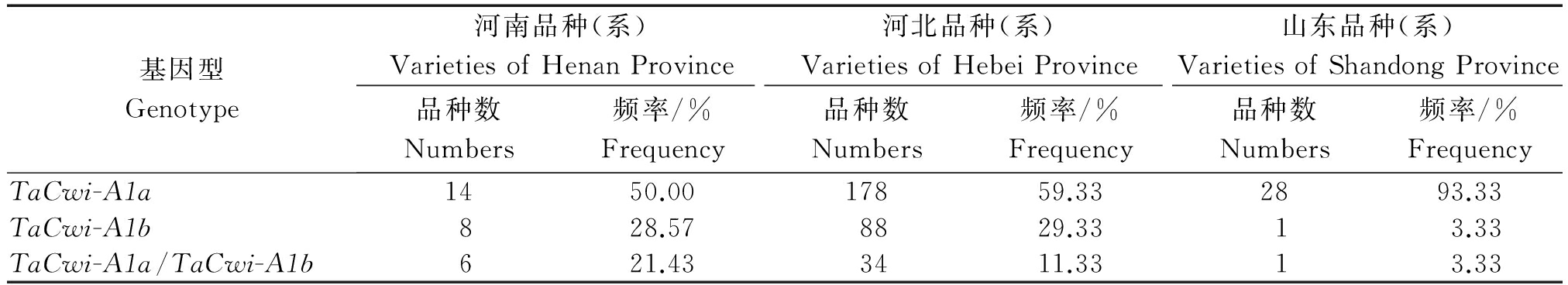
基因型Genotype河南品种(系)VarietiesofHenanProvince河北品种(系)VarietiesofHebeiProvince山东品种(系)VarietiesofShandongProvince品种数Numbers频率/%Frequency品种数Numbers频率/%Frequency品种数Numbers频率/%FrequencyTaCwi-A1a1450.0017859.332893.33TaCwi-A1b828.578829.3313.33TaCwi-A1a/TaCwi-A1b621.433411.3313.33
表5 参加区试小麦品种资源中TaCwi-A1a和TaCwi-A1b在各环境下的千粒质量
Tab.5 The TKW of region test varieties with TaCwi-A1a and TaCwi-A1b in three environments
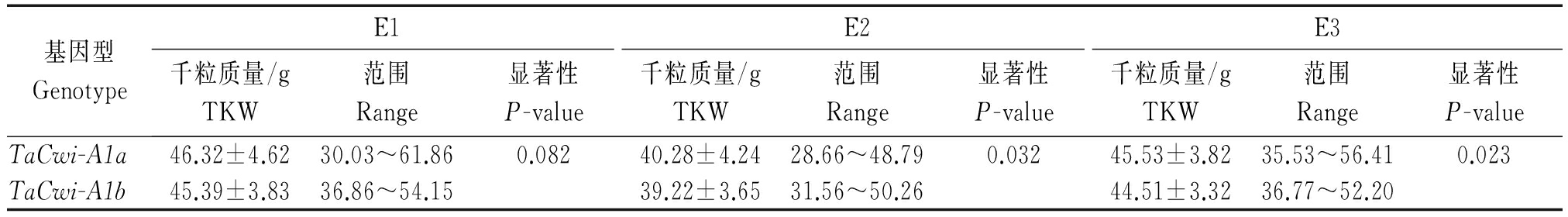
基因型GenotypeE1E2E3千粒质量/gTKW范围Range显著性P-value千粒质量/gTKW范围Range显著性P-value千粒质量/gTKW范围Range显著性P-valueTaCwi-A1a46.32±4.6230.03~61.860.08240.28±4.2428.66~48.790.03245.53±3.8235.53~56.410.023TaCwi-A1b45.39±3.8336.86~54.1539.22±3.6531.56~50.2644.51±3.3236.77~52.20
对非杂合型的102份自育材料中2种等位变异类型在不同环境条件下的千粒质量表型值进行了分析(表6),t检验表明,E1、E2、E3 3个环境下,TaCwi-A1a基因型材料的千粒质量均显著高于TaCwi-A1b(E1的P值<0.05,E2的P值<0.001,E3的P值<0.05),平均千粒质量分别高3.16,4.80,2.47 g。
表6 自育品系中TaCwi-A1a和TaCwi-A1b在各环境下的千粒质量
Tab.6 The TKW of breeding varieties with TaCwi-A1a and TaCwi-A1b in three environments
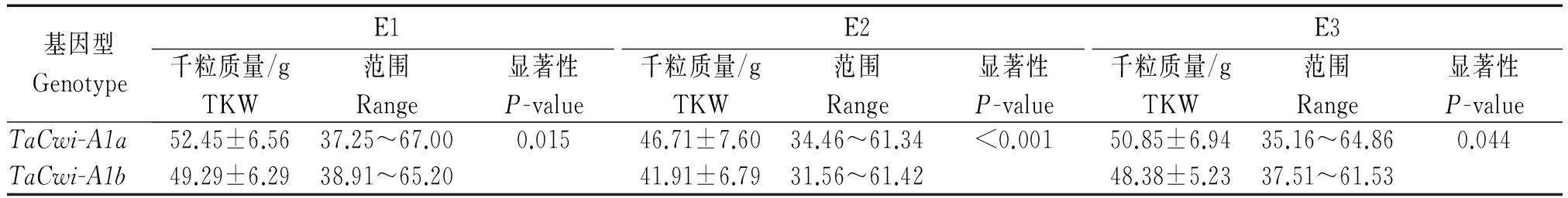
基因型GenotypeE1E2E3千粒质量/gTKW范围Range显著性P-value千粒质量/gTKW范围Range显著性P-value千粒质量/gTKW范围Range显著性P-valueTaCwi-A1a52.45±6.5637.25~67.000.01546.71±7.6034.46~61.34<0.00150.85±6.9435.16~64.860.044TaCwi-A1b49.29±6.2938.91~65.2041.91±6.7931.56~61.4248.38±5.2337.51~61.53
以E3环境下的表型数据为基础,对上述供试材料中TaCwi-A1等位变异与粒重各构成要素的差异性进行了分析(表7),结果表明,TaCwi-A1a基因型材料与TaCwi-A1b基因型材料的粒长(P<0.05)和粒宽(P<0.05)差异达到显著水平。在粒重及其构成要素中,增幅顺序为:千粒质量(5.11%)>粒长(3.51%)>籽粒面积(2.74%)>粒宽(2.37%)。上述分析同样证明,TaCwi-A1a对粒重及其构成要素是优异的等位变异。
表7 自育品系中TaCwi-A1等位变异的粒重表型性状比较
Tab.7 Comparison of grain phenotypic traits of TaCwi-A1 polymorphism in breeding varieties
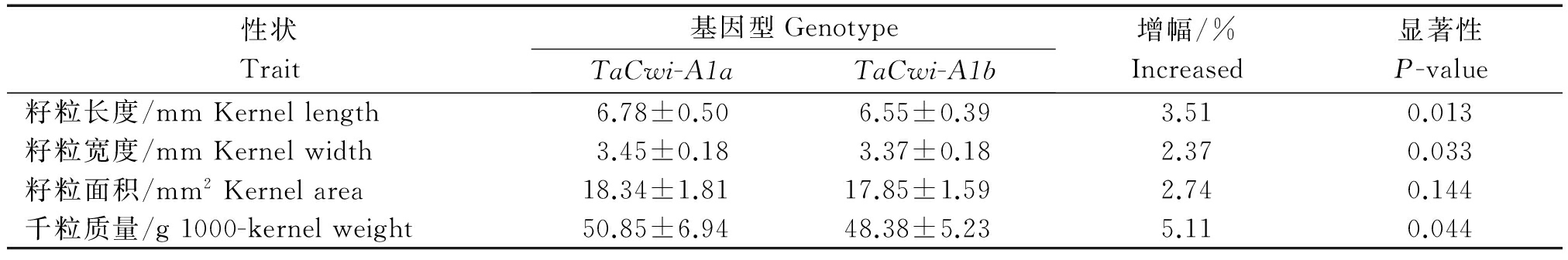
性状Trait基因型GenotypeTaCwi-A1aTaCwi-A1b增幅/%Increased显著性P-value籽粒长度/mmKernellength6.78±0.506.55±0.393.510.013籽粒宽度/mmKernelwidth3.45±0.183.37±0.182.370.033籽粒面积/mm2Kernelarea18.34±1.8117.85±1.592.740.144千粒质量/g1000-kernelweight50.85±6.9448.38±5.235.110.044
冀中南小麦育种以多穗型品种为主,粒重提升潜力较大。本研究发现,自育品系中粒重优异等位变异TaCwi-A1a的分布频率明显低于河北省的平均分布频率,今后我们应在亲本选配方面注意引入高粒重的TaCwi-A1a供体。
3 讨论与结论
TaCwi-A1基因在光合产物运输过程中起重要作用,通过对籽粒灌浆速率起调控作用,进而影响籽粒千粒质量大小。利用豆麦/石4185 F2:3群体对TaCwi-A1基因进行QTL分析,可以解释粒重表型变异的4.8%,具有TaCwi-A1a基因型的品种有较高的千粒质量,而具有TaCwi-A1b基因型的品种通常千粒质量较低[4]。本研究中对黄淮麦区小麦品种资源的TaCwi-A1功能标记检测结果表明,TaCwi-A1a的分布频率为65.03%明显高于TaCwi-A1b的分布频率34.97%。据笔者了解,绝大多数育种单位并没有开展分子标记辅助选择的工作,育种家们通过田间农艺性状人为选择,很自然的保留了TaCwi-A1a基因型的材料,证明TaCwi-A1a基因型确实为粒重优异的等位变异。
目前小麦千粒质量研究所用的群体多为回交群体、DH群体和重组自交系等[18-20],这些群体往往是基于粒重性状差异很大的2个亲本杂交衍生形成的,由于遗传背景狭窄,难以体现特定基因在不同环境条件、不同遗传背景下的真实情况。本研究利用审定品种和区试品系等贴近生产实际的自然群体作为研究对象,获得了更为广泛的遗传基础,更有利于评价基因不同等位变异与性状的关联分析。此外,考虑到小麦千粒质量受气候、生产条件等因素影响,年际间差异较大。笔者获取了连续2个生长周期共3个环境下的表型数据,希望通过优化遗传群体类型和多年多点试验种植,更准确评价粒重基因的分子标记辅助选择应用价值。
由于小麦粒重是由粒长、粒宽、粒形、质地等要素构成的,虽然各要素对粒重整体性状的“贡献率”有所差异,但它们与粒重间均呈正相关,且各构成要素之间无负相关[21-22]。因此,通过遗传改良小麦粒重的任一构成要素均可达到提高小麦千粒质量的目的。本研究中对于审定品种和自育品系进行了一个环境下的数据分析,对于各要素与粒重和环境因子的相关性分析还需要积累更多环境数据,有待于下一步深入研究。
河北省农林科学院遗传生理研究所选育材料的千粒质量明显高于其他单位的,无论是TaCwi-A1a基因型,还是TaCwi-A1b基因型,看来本研究的自育品系可能携带着更重要的控制高千粒质量(或大粒)的基因,有待进一步挖掘与解析。
小麦粒重基因TaCwi-A1 2种等位变异在黄淮麦区品种资源中分布频率呈现TaCwi-A1a明显高于TaCwi-A1b。TaCwi-A1a基因型材料与TaCwi-A1b基因型材料的千粒质量、粒长、粒宽和籽粒面积差异都达到显著水平,且都有不同程度的增效,综合而言,TaCwi-A1a基因型对应更高的籽粒表型性状数据,对粒重及其构成要素是优异的等位变异。总之,TaCwi-A1可用于小麦粒重的分子标记辅助选择。
参考文献:
[1] 林作楫,揭声慧,雷振生,等.近60年黄淮麦区冬小麦育种规律演变研究[J].现代农业科技,2011(24):103-105,108.
[2] 庄巧生.中国小麦品种改良及系谱分析[M].北京:中国农业出版社,2003:497-518.
[3] Wang L,Ge H,Hao C,et al.Identifying loci influencing 1,000-kernel weight in wheat by microsatellite screening for evidence of selection during breeding[J].PLoS One,2012,7(2):e29432.
[4] Ma D Y,Yan J,He Z H,et al.Characterization of a cell wall invertase gene TaCwi-A1 on common wheat chromosome 2A and development of functional markers[J].Molecular Breeding,2012,29(1):43-52.
[5] Jiang Q,Hou J,Hao C,et al.The wheat(T.aestivum)sucrose synthase 2 gene(TaSus2)active in endosperm development is associated with yield traits[J].Functional & Integrative Genomics,2011,11(1):49-61.
[6] Su Z,Hao C,Wang L,et al.Identification and development of a functional marker of TaGW2 associated with grain weight in bread wheat(Triticum aestivum L.)[J].Theoretical and Applied Genetics,2011,122(1):211-223.
[7] Yang Z,Bai Z,Li X,et al.SNP identification and allelic-specific PCR markers development for TaGW2,a gene linked to wheat kernel weight[J].Theoretical and Applied Genetics,2012,125(5):1057-1068.
[8] Qin L,Hao C,Hou J,et al.Homologous haplotypes,expression,genetic effects and geographic distribution of the wheat yield gene TaGW2[J].BMC Plant Biology,2014,14:107.
[9] Zhang L,Zhao Y,Gao L,et al.TaCKX6-D1,the ortholog of rice OsCKX2,is associated with grain weight in hexaploid wheat[J].The New Phytologist,2012,195(3):574-584.
[10] Chang J,Zhang J,Mao X,et al.Polymorphism of TaSAP1-A1 and its association with agronomic traits in wheat[J].Planta,2013,237(6):1495-1508.
[11] Guo Y,Sun J J,Zhang G Z,et al.Haplotype,molecular marker and phenotype effects associated with mineral nutrient and grain size traits of TaGS1a in wheat[J].Field Crops Research,2013,154(3):119-125.
[12] Zhang Y J,Liu J D,Xia X C,et al.TaGS-D1,an ortholog of rice OsGS3,is associated with grain weight and grain length in common wheat[J].Molecular Breeding,2014,34(3):1097-1107.
[13] Dong L L,Wang F M,Liu T,et al.Natural variation of TaGASR7-A1 affects grain length in common wheat under multiple cultivation conditions[J].Molecular Breeding,2014,34(3):937-947.
[14] 韩利明,杨芳萍,夏先春,等.株高、粒重及抗病相关基因在不同国家小麦品种中的分布[J].麦类作物学报,2011,31(5):824-831.
[15] 相吉山,穆培源,桑 伟,等.小麦粒重基因TaCwi-A1功能标记CWI22、CWI21的验证及应用[J].中国农业科学,2014,47(13):2671-2709.
[16] 张祖建,郎有忠,潘美红,等.粳稻小粒穗的症状及其籽粒性状的变化特征[J].中国农业科学,2006,39(8):1536-1544.
[17] 王国英.基因工程实验技术[M].北京:中国农业科技出版,1997:43-46.
[18] Huang X,Kempf H,Ganal M,et al.Advanced backcross QTL analysis in progenies derived from a cross between a German elite winter wheat variety and a synthetic wheat(Triticum aestivum L.)[J].Theoretical and Applied Genetics,2004,109(5):933-943.
[19] Mccartney C A,Somers D J,Humphreys D G,et al.Mapping quantitative trait loci controlling agronomic traits in the spring wheat cross RL4452 ×AC Domain[J].Genome,2005,48(5):870-883.
[20] Wang R,Hai L,Zhang X,et al.QTL mapping for grain filling rate and yield-related traits in RILs of the Chinese winter wheat population Heshangmai×Yu8679[J].Theoretical and Applied Genetics,2009,118(2):313-325.
[21] 常 成,张海萍,张秀英,等.小麦PEBP-like基因等位变异与籽粒大小、粒重关系研究[J].分子植物育种,2009,7(1):23-27.
[22] Sun X Y,Wu K,Zhao Y,et al.QTL analysis of kernel shape and weight using recombinant inbred lines in wheat[J].Euphytica,2008,165(3):615-624.
《华北农学报》征订启事
《华北农学报》1986年创刊,由河北、北京、天津、河南、山西、内蒙古六省市区农科院、农学会联合主办,为全国首家跨省、市、区多单位联办的农业学术刊物。本刊立足华北,面向全国和全世界。主要刊载农作物、果树、水产、畜牧、资环、植保等农业基础学科的原创性研究论文、专论、综述、研究简报等,报道农业学术动态。主要服务于农业高等院校师生和农业科研机构的研究人员。
《华北农学报》为中国科学引文数据库核心期刊(CSCD核心库)、中文核心期刊、中国科技核心期刊、RCCSE中国核心学术期刊和中国农业核心期刊。在2014年版《中文核心期刊要目总览》综合性农业科学类核心期刊中排名第2位,为我国有影响力的农业学术刊物。《华北农学报》多次荣获国家级及省级奖励:全国优秀科技期刊评比三等奖、全国优秀农业期刊学术类一等奖、首届华北优秀期刊、首届北方十佳期刊、中国北方优秀期刊、河北省荣誉期刊、河北省精品期刊、河北省十佳期刊及河北省优秀期刊等奖项;2011年被评选为“中国精品科技期刊”。
《华北农学报》国内外公开发行, 国内统一刊号:CN13-1101/S,国际刊号ISSN 1000-7091。双月刊,双月28日出版,国际标准大16开本,240页,每期定价12元,全年72.00元。邮发代号: 18-10,国外发行代号:5918。全国各地邮局均可订阅。可随时汇款到编辑部订阅,请注明刊名、份数、姓名、地址、邮编及电话。
欢迎订阅、欢迎投稿。
地址:石家庄市和平西路598号《华北农学报》编辑部
邮编:050051
电话:0311-87652166
E-mail:hbnxb@163.com
网址:http://www.hbnxb.net/
Functional Analysis and Distribution of Allelic Variations of TaCwi-A1 Gene Related to Kernel Weight in Yellow and Huai River Valleys Facultative Wheat Zone
LIU Yongwei1,ZHOU Shuo1,WANG Xuezheng2,SUN Guozhong1,ZHU Jinyong1,HAN Qiufen1,LI Chunjie3,ZHAO He1,WANG Haibo1
(1.Institute of Genetics and Physiology,Hebei Academy of Agriculture and Forestry Sciences,Plant Genetic Engineering Center of Hebei Province,Shijiazhuang 050051,China;2.Dryland Farming Institute,Hebei Academy of Agricultural and Forestry Science,Hengshui 053000,China;3.The Seed Management Station of Hebei Province,Shijiazhuang 050031,China)
Abstract:In order to evaluate the breeding application value of two kind of allelic variations(TaCwi-A1a and TaCwi-A1b)of TaCwi-A1 gene,which was related to wheat kernel weight.In this study,539 wheat varieties(lines)in Yellow and Huai River Valleys Facultative Wheat Zone in China were genotyped using functional markers of TaCwi-A1 to determine the allelic variation types,in which the frequencies of TaCwi-A1a and TaCwi-A1b could be detected.To compare the differences of phenotypic traits of TaCwi-A1a and TaCwi-A1b,thousand kernel weight(TKW)of released varieties,region test varieties,and breeding varieties was evaluated in three environments;and kernel length(KL),kernel width(KW),and kernel area(KA)was also evaluated in one environment.The results showed that the frequencies of TaCwi-A1a and TaCwi-A1b were 65.03% and 34.97% in all test materials,in which the frequency of TaCwi-A1a was significantly higher than TaCwi-A1b.Analysis of phenotypic traits of grain showed that TKW of TaCwi-A1a was significantly higher than TaCwi-A1b in released varieties,region test varieties and breeding varieties.Similarly,KL and KW of TaCwi-A1a were also significantly higher than TaCwi-A1b.This study further verified the additive effect of TaCwi-A1a on phenotypic traits of grain,which showed that TaCwi-A1a was superior allelic variation for the grain weight and its constituent elements.In addition,the study identified the distribution of TaCwi-A1 in Yellow and Huai River Valleys Facultative Wheat Zone in China,and provided useful information for the marker-assisted selection of wheat.
Key words:Common wheat;TaCwi-A1;Molecular marker;Allelic variation;Thousand kernel weight;Kernel length;Kernel width
收稿日期:2017-01-23
基金项目:河北省农林科学院科学技术研究与发展计划项目(A2015110102);河北省现代农业创新工程项目(2017038997);北京全式金生物技术有限公司Trans助研梦想基金(Trans-RasDF-003)
作者简介:刘永伟(1981-),男,内蒙古乌拉特中旗人,助理研究员,硕士,主要从事作物分子育种研究。
通讯作者:赵 和(1962-),男,河北康保人,研究员,硕士,主要从事小麦分子育种研究。 王海波(1958-),男,河北满城人,研究员,博士,博士生导师,主要从事作物遗传与生物技术研究。
中图分类号:S512.01
文献标识码:A
文章编号:1000-7091(2017)02-0131-07
doi:10.7668/hbnxb.2017.02.020