

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
外施2,4-表油菜素内酯缓解冬小麦拔节期冻害胁迫效应
引用本文
张奥深, 雍晓宇, 韩巧霞, 姚永伟, 师焕婷, 李鸽子, 康国章. 外施2,4-表油菜素内酯缓解冬小麦拔节期冻害胁迫效应[J].中国地质调查, 2022,37(2): 78-83.
ZHANG Aoshen, YONG Xiaoyu, HAN Qiaoxia, YAO Yongwei, SHI Huanting, LI Gezi, KANG Guozhang. Alleviating Effect of External Application of 2,4-Epbrassinolide on Freezing Stress in Winter Wheat at Jointing Stage[J].Geological Survey of China, 2022,37(2): 78-83.
doi: 10.7668/hbnxb.20192445
Permissions
ZHANG Aoshen, YONG Xiaoyu, HAN Qiaoxia, YAO Yongwei, SHI Huanting, LI Gezi, KANG Guozhang. Alleviating Effect of External Application of 2,4-Epbrassinolide on Freezing Stress in Winter Wheat at Jointing Stage[J].Geological Survey of China, 2022,37(2): 78-83.
doi: 10.7668/hbnxb.20192445
Copyright@2022, 《华北农学报》编辑部
版权所有 © 2022 《华北农学报》编辑部
外施2,4-表油菜素内酯缓解冬小麦拔节期冻害胁迫效应
作者简介:张奥深(1997—),男,河南驻马店人,在读硕士,主要从事作物逆境生理与分子生物学研究。
摘要
为探讨外源油菜素内酯对冻害胁迫条件下拔节期小麦生理特性、转录水平及生长的影响,选用半冬性小麦品种百农207为材料,采用0.1 mg/L的2,4-表油菜素内酯(EBR)浇施拔节期小麦,研究冻害胁迫下小麦叶绿素含量(SPAD值)、丙二醛(MDA)含量、相对电导率、游离脯氨酸(Pro)含量、抗逆相关基因表达量、幼穗受冻率、籽粒产量、千粒质量及干物质质量的变化。结果表明,冻害胁迫前,对照组与处理组的SPAD值、MDA含量和相对电导率无显著差异,而Pro含量处理组显著低于对照组;在冻害胁迫后,与对照相比,外施EBR显著降低了小麦叶片的MDA含量和相对电导率,提高了SPAD值和Pro含量。冻害胁迫48 h,处理组小麦植株叶片抗逆相关基因 SOD、 POD、 CAT、 P5CS和 WCS120的相对表达量均显著高于对照;冻害胁迫后,与对照组相比,处理组植株幼穗受冻率显著降低,比对照降低了20.93百分点,而籽粒产量、千粒质量和干物质质量显著增加,分别增加了19.38,2.37,54.40 g。综上,小麦拔节期外施EBR可以通过降低叶片MDA含量,提高Pro含量、叶绿素含量以及抗逆相关基因 SOD、 POD、 CAT、 P5CS和 WCS120的表达量,从而提高小麦的抗冻性,减轻低温冻害对冬小麦造成的产量损失。
关键词:
冬小麦; 2,4-表油菜素内酯; 冻害胁迫; 拔节期
中图分类号:S512.01
文献标志码:A
文章编号:1000-7091(2022)02-0078-06
Alleviating Effect of External Application of 2,4-Epbrassinolide on Freezing Stress in Winter Wheat at Jointing Stage
Abstract
In order to investigate the effects of exogenous brassinolide on physiological characteristics,transcriptional level and growth of wheat at jointing stage under freezing stress,the semi-winter wheat variety Bainong 207 was used as material,and irrigated with 0.1 mg/L 2,4-Epibrassinolide(EBR)to study its chlorophyll content(SPAD value),malondialdehyde(MDA)content,relative conductivity,free proline(Pro)content,stress resistance related gene expression,young panicle freezing rate,grain yield,1 000 grains weight and dry matter quality.The results showed that before freezing stress,there was no significant difference in SPAD value,MDA content and relative conductivity between the control group and the treatment group,but the Pro content in the treatment group was significantly lower than that in the control group;after freezing stress,compared with the control,external application of EBR significantly reduced the MDA content and relative conductivity of wheat leaves,and increased the SPAD value and Pro content.At 48 hours of freezing stress,the relative expression of stress resistance related genes SOD, POD, CAT, P5CS,and WCS120 in the treatment group were significantly higher than those in the control.After freezing stress,compared with the control,the freezing rate of young panicles in the treatment group decreased significantly by 20.93 percentage point,while the grains yield,1 000 grain weight and dry matter mass increased significantly by 19.38,2.37,54.40 g,respectively.In conclusion,external application of EBR at jointing stage of wheat can improve the frost resistance of wheat and reduce the yield loss of winter wheat caused by low temperature and freezing injury by reducing the content of MDA,increasing the content of Pro,chlorophyll and the expression of stress resistance related genes SOD, POD, CAT, P5CS,and WCS120.
Keyword:
Winter wheat; 2,4-Epbrassinolide; Freezing stress; Jointing stage
小麦作为我国三大粮食作物之一, 其产业发展直接影响到国家粮食安全和社会稳定。近年来, 随着全球气候变暖, 极端低温灾害频繁发生, 严重影响了小麦的稳产、高产[1], 其中倒春寒是黄淮麦区最严重的灾害, 在我国冬麦主产区黄淮麦区, 小麦冻害发生的频率越来越高, 对小麦造成的危害也越来越严重[2]。小麦拔节期到抽穗前是小麦生长速度最快, 生长量最大的时期, 叶面积及茎穗的长度和体积成倍或几十倍增长, 干物质积累也进入迅速增长阶段, 此时小麦生长旺盛, 抗寒能力弱[3], 低温对小麦造成的不利影响较为严重[4]。
油菜素内酯(Brassinolide, BR)又称芸薹素内酯[5], 是广泛存在于植物体内、参与各种生理活动的一种天然化合物, 被称为第六种植物激素[6]。BR是公认的高效、广谱、无毒的一种植物生长调节剂。2, 4-表油菜素内酯(2, 4-Epibrassinolide, EBR)是一种人工合成的高活性油菜素内酯类似物, 外施EBR能够提高作物的抗逆能力[7]。尚宏芹等[8]研究表明, 叶面喷施EBR可通过提高小麦幼苗的抗氧化酶活性和抗氧化物质含量、降低MDA和H2O2含量, 减轻重金属汞对幼苗的伤害, 促进小麦幼苗的生长。孙玉珺等[9]研究表明, 外源BR处理可提高幼苗生物量、抗氧化酶活性以及Pro、可溶性糖和可溶性蛋白含量, 降低MDA含量, 缓解低温胁迫对玉米生长的抑制程度。刘丽杰等[10]研究发现, 利用0.1 mg/L的EBR处理小麦幼苗, 可提高小麦的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性, 可溶性糖和Pro含量显著提高, 降低MDA含量, 从而提高小麦的抗冻性。前人从生理层面对EBR提高作物的耐受性进行研究, 证实EBR对多种逆境胁迫具有缓解作用, 但从基因层面对EBR的作用机制研究较少。
本试验以半冬性品种百农207为材料, 在小麦拔节期外源浇施EBR, 并利用人工气候室模拟低温冻害, 研究冻害胁迫条件下小麦植株生理指标、抗逆相关基因表达量、幼穗受冻率、籽粒产量、千粒质量以及地上部干物质质量的变化, 探究外源EBR缓解小麦低温冻害的生理机制, 以期为减轻倒春寒对小麦的危害提供理论依据和技术支撑。
1 材料和方法
1.1 试验材料与设计
供试材料为普通半冬性小麦品种百农207。试验采用盆栽, 2020年秋季种植在河南省郑州市毛庄河南农业大学科技园试验田中, 盆的直径为30 cm, 高为30 cm, 三叶期每盆定苗16株。盆栽用土为0~30 cm的耕层土壤, 每盆装过筛壤土8 kg, 并均匀施入尿素2.3 g、磷肥0.8 g和钾肥3.4 g, 之后埋于试验田中, 盆的顶部边缘与地面齐平。
在前期试验筛选EBR浓度的基础上, 待小麦生长至拔节期时浇施浓度为0.1 mg/L的EBR 400 mL, 对照(CK)施用同等体积的水。小麦植株充分吸收24 h后, 将盆栽移入人工气候室内-5 ℃低温连续处理48 h, 在处理0, 24, 48 h分别剪取主茎最上部全展叶, 用于生理指标的测定及基因表达分析, 每个处理重复3次。
1.2 生理指标测定
在冻害胁迫处理前后利用SPAD-502叶绿素仪测定小麦最上部全展叶的SPAD值, 每盆测5片叶片取平均值。利用硫代巴比妥酸(TCA-TBA)显色法测定叶片MDA含量, 采用酸性茚三酮法测定Pro含量, 利用电导率仪测定叶片相对电导率[11]。
1.3 基因表达分析
取小麦叶片0.1 g, 利用总RNA提取试剂盒(Vazyme, 南京)提取总RNA, 反转录试剂盒(Vazyme, 南京)合成cDNA第一链。在前期研究中, 小麦高产抗逆分子调控创新团队筛选出与冻害胁迫相关的5个抗逆基因:SOD[12]、POD[13]、CAT[14]、P5CS[15]和WCS120[16]。本研究以小麦Actin作为内参基因, 对5个抗逆相关基因进行转录水平的检测。采用软件Primer Premier 5.0设计引物(表1), 由河南尚亚生物技术有限公司进行合成。2-Δ Δ Ct法计算基因的相对表达量。
1.4 小麦幼穗受冻率调查和籽粒产量、千粒质量、干物质质量测定
在拔节期对盆栽小麦外源施用EBR 24 h后, 进行冻害胁迫处理(48 h), 自然条件下恢复生长, 15 d后调查幼穗受冻后死亡情况, 取其平均值计算幼穗
| 表1 实时荧光定量PCR引物信息 Tab.1 qRT-PCR primer information |
受冻率; 于成熟期按盆收获籽粒计算籽粒产量; 按处理随机选取1 000个籽粒确定千粒质量; 将收获后小麦植株地上部放入烘箱, 105 ℃杀青30 min后80 ℃烘干, 计算干物质质量。
1.5 数据处理
采用Microsoft Excel 2010进行数据处理, 采用SPSS 22.0软件进行方差分析, 采用Origin 9.0作图。
2 结果与分析
2.1 小麦拔节期外施EBR冻害胁迫后植株叶片SPAD值的变化
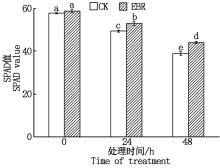
如图1所示, 冻害胁迫前, 外施EBR处理组与对照的SPAD值无显著差异, 在冻害胁迫后, 叶片的SPAD值随冻害胁迫时间的延长呈下降趋势, 在同一胁迫时间处理组的SPAD值始终显著高于对照, 这表明小麦拔节期外施EBR可以缓解冻害胁迫下叶片衰老, 延长叶片的功能期。
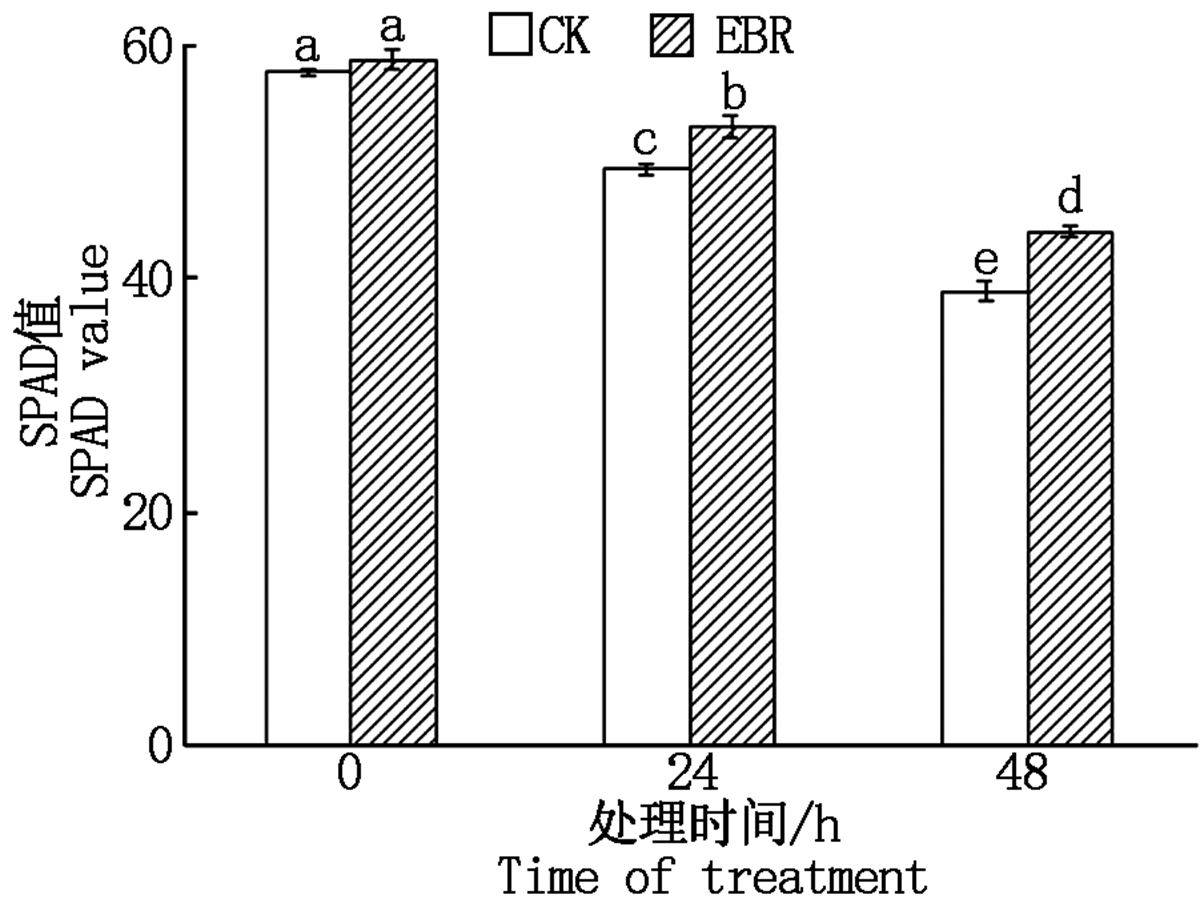 | 图1 小麦拔节期外施EBR冻害胁迫后叶片的SPAD值 柱上不同小写字母均表示在5%水平差异显著。图2— 5同。Fig.1 SPAD value in wheat leaves under freezing stress after external application of EBR at jointing stage The different lowercase letters on the columns indicate significant differences at the 5% level.The same as Fig.2— 5. |
2.2 小麦拔节期外施EBR冻害胁迫后植株叶片MDA含量和相对电导率的变化
冻害胁迫处理前外施EBR的小麦植株和对照植株叶片MDA含量差异不显著; 冻害胁迫24 h对照组MDA含量显著升高, 而外施EBR处理组小麦植株叶片MDA含量与冻害胁迫前无显著变化; 而冻害胁迫处理时间达到48 h, 二者的MDA含量均较处理24 h显著下降, 且处理组MDA含量显著低于对照组。结果表明, 拔节期外施EBR可以降低小麦植株叶片MDA含量, 从而减轻其膜脂过氧化程度(图2)。
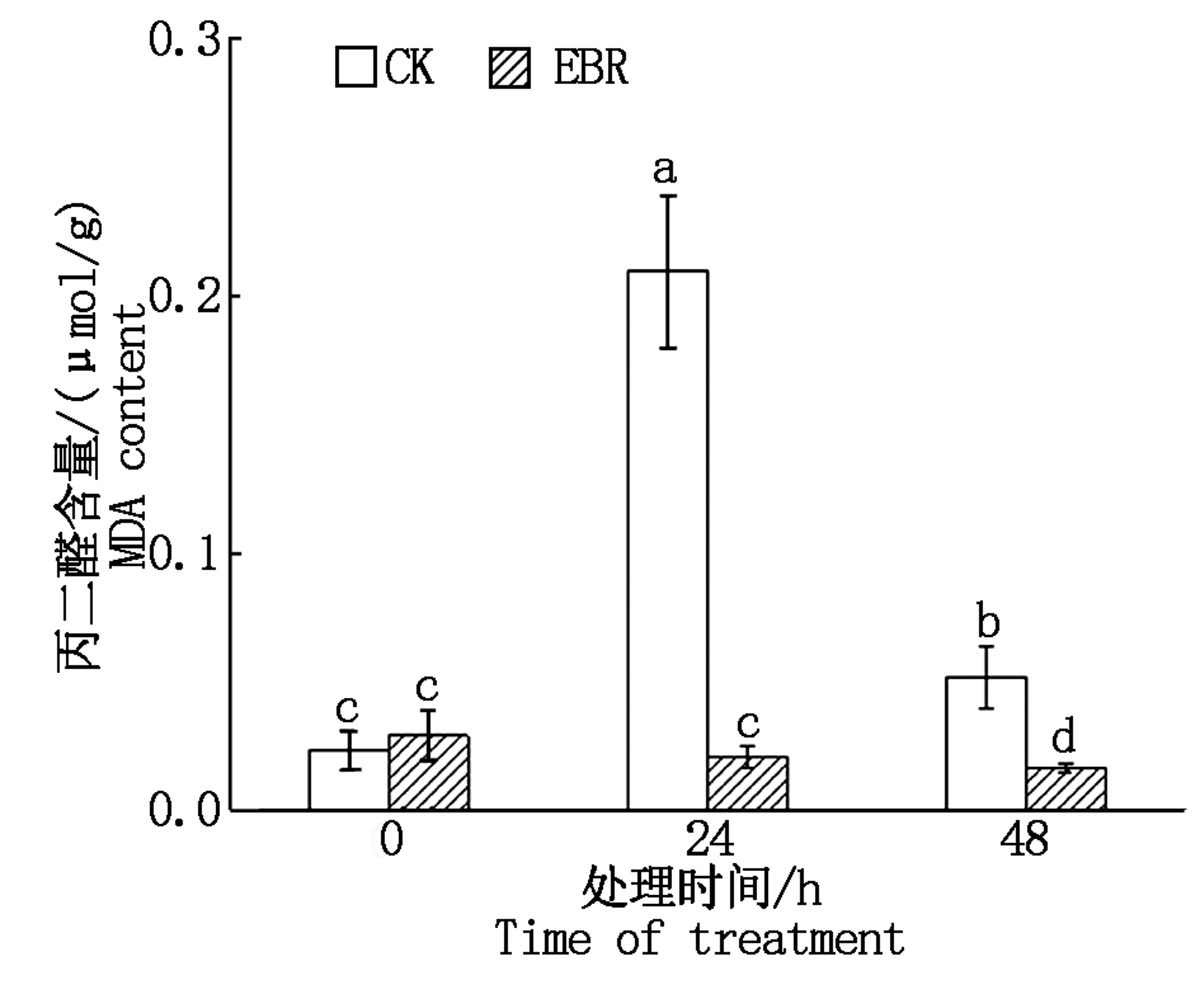 | 图2 小麦拔节期外施EBR冻害胁迫后叶片的MDA含量Fig.2 MDA content in wheat leaves under freezing stress after external application of EBR at jointing stage |
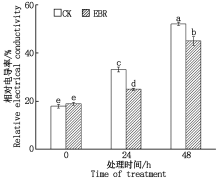
在冻害胁迫处理前外施EBR植株叶片的相对电导率与对照无显著差异, 在低温胁迫24, 48 h, 外施EBR的植株相对电导率均显著低于对照(P< 0.05), 分别降低了8.19, 7.03百分点(图3)。
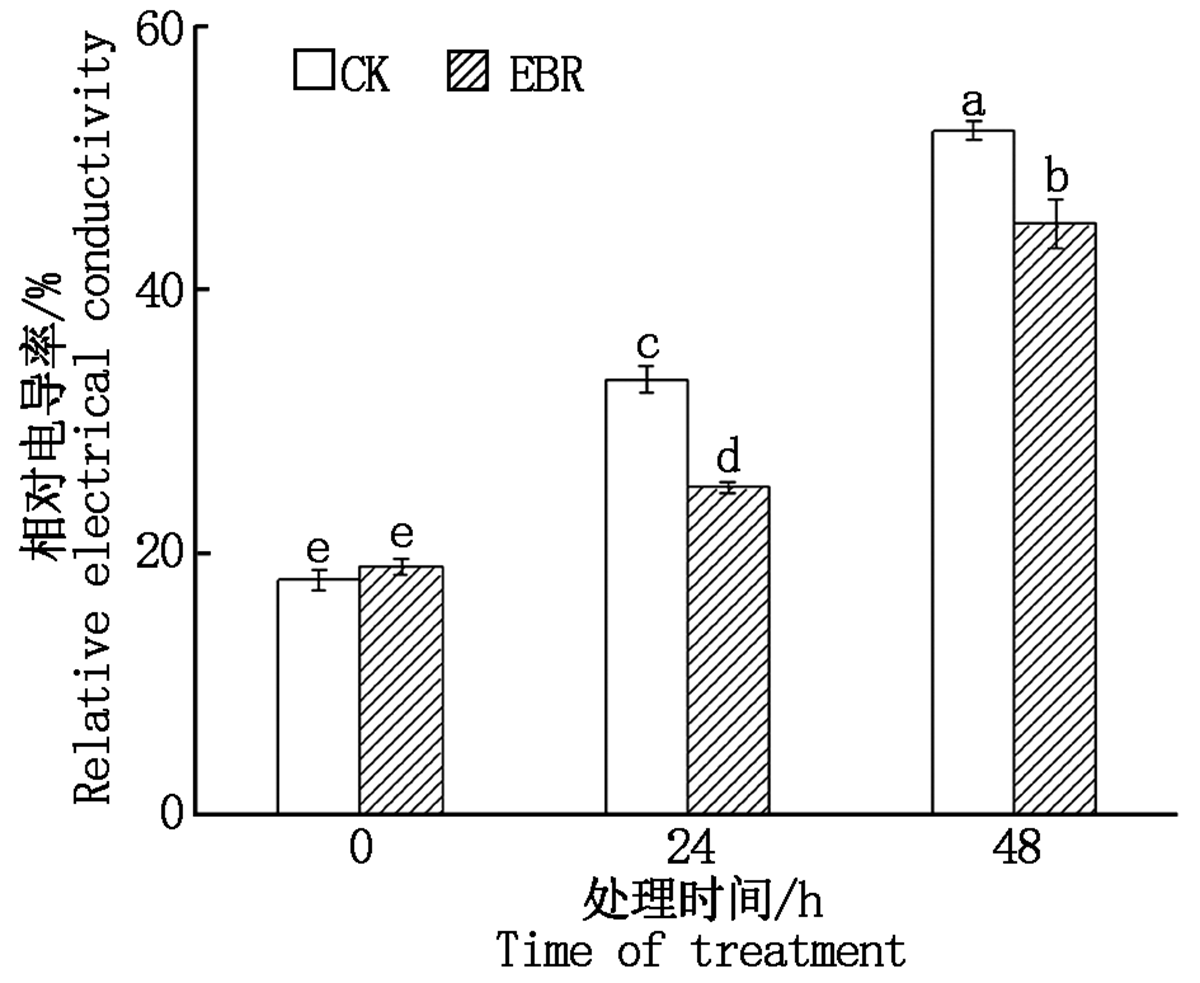 | 图3 小麦拔节期外施EBR冻害胁迫后叶片的相对电导率Fig.3 Relative conductivities in wheat leaves under freezing stress after external application of EBR at jointing stage |
2.3 小麦拔节期外施EBR冻害胁迫后植株叶片Pro含量的变化
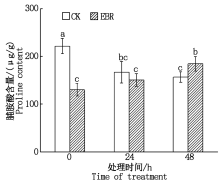
由图4可知, 对照组在冻害胁迫下, Pro含量呈现逐渐下降趋势; 而外施EBR的处理组在胁迫条件下, 叶片的Pro含量逐渐上升; 在冻害胁迫48 h, 处理组小麦叶片的Pro含量显著高于对照组(P< 0.05)。
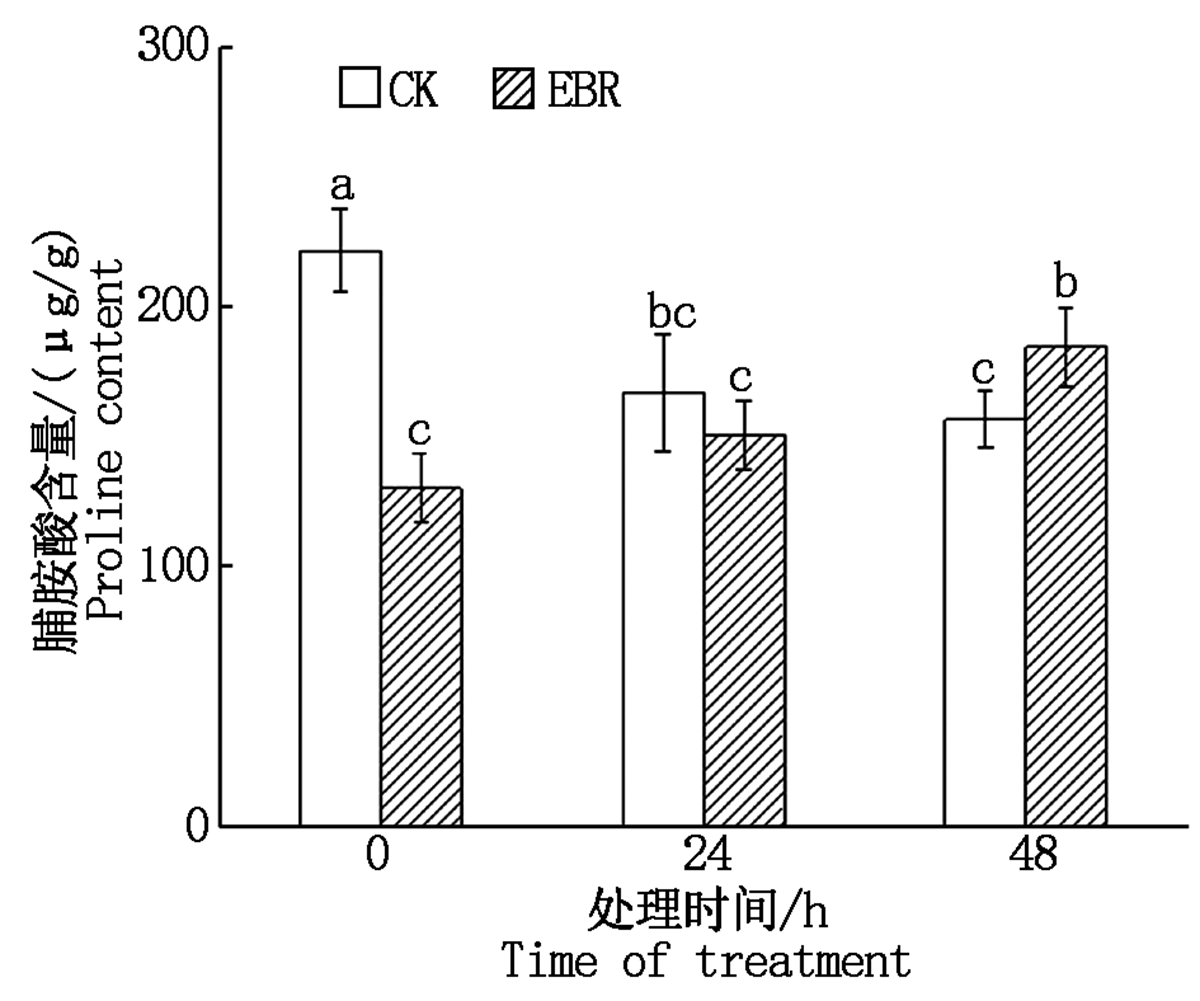 | 图4 小麦拔节期外施EBR冻害胁迫后叶片Pro含量Fig.4 Proline contents in wheat leaves under freezing stress after external application of EBR at jointing stage |
2.4 小麦拔节期外施EBR冻害胁迫后植株叶片抗逆相关基因表达量变化
在冻害胁迫处理前, 除P5CS其余4个抗逆相关基因的表达量处理组均显著高于对照组(P< 0.05); 处理组小麦植株在冻害胁迫24 h, 叶片中的SOD、POD和CAT的表达量较胁迫处理前均显著降低, 而P5CS和WCS120的表达量均显著上升; 对照组在冻害胁迫处理24 h, SOD、CAT的表达量较冻害胁迫处理前无显著变化, P5CS和WCS120的表达量均显著上升。冻害胁迫处理48 h与24 h相比, 处理组的SOD、POD、CAT和WCS120的相对表达量均显著提高; 而对照组的SOD、P5CS和WCS120表达量无显著变化。在冻害胁迫处理48 h, 处理组的5个抗逆相关基因表达量均显著高于对照(P< 0.05)(图5)。
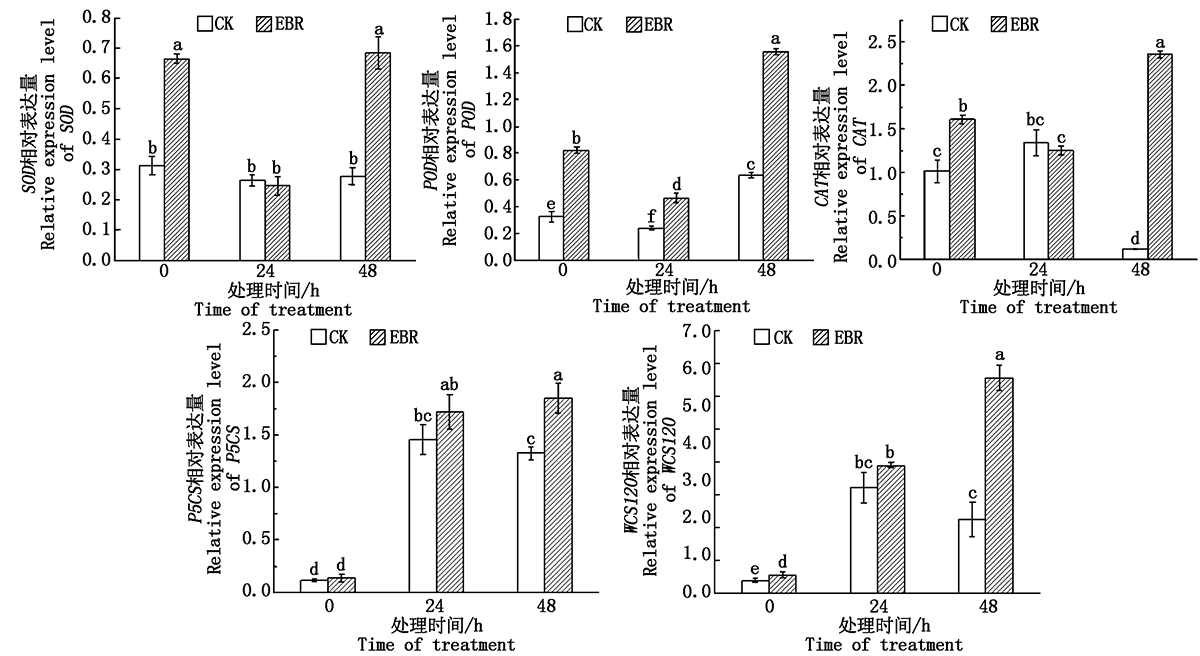 | 图5 小麦拔节期外施EBR冻害胁迫后叶片抗逆相关基因的相对表达量Fig.5 Relative expression leaves of stress resistance related genes in wheat leaves under freezing stress after external application of EBR at jointing stage |
2.5 小麦拔节期外施EBR冻害胁迫后植株幼穗受冻率、籽粒产量、千粒质量和干物质质量的变化
外施EBR的处理组小麦植株幼穗受冻率比对照组降低了20.93百分点; 处理组的植株籽粒产量和干物质质量每盆分别较对照增加了19.38, 54.40 g, 处理组千粒质量较对照增加了2.37 g, 均显著高于对照。说明在拔节期外施EBR能够提高小麦幼穗的抗冻性和干物质积累量, 从而提高小麦产量(表2)。
| 表2 小麦拔节期外施EBR冻害胁迫后幼穗受冻率、籽粒产量、千粒质量和干物质质量 Tab.2 The damage rates of young ears, grain yields, 1 000 grains weights and dry matter qualities in wheat under freezing stress after external application of EBR at jointing stage |
3 结论与讨论
油菜素内酯是一种多羟基甾类化合物, 是调节植物逆境胁迫伤害的重要生长调节剂, 参与不同植物的生长、发育和抵抗外界胁迫等生物学功能[17, 18]。外源EBR对植物逆境胁迫缓解方面的生理作用日益受到关注[19, 20, 21]。华智锐等[22]研究发现, 在干旱胁迫下, 经BR处理的小麦幼苗抗氧化酶活性提高, MDA含量降低, 相对含水量提高, 从而提高了小麦幼苗的抗旱性。李淑叶[23]在低温处理前对棉花叶面分别喷施不同浓度的EBR和蒸馏水, 结果表明, 外源喷施适宜浓度的EBR较喷施蒸馏水可显著提高叶片Pro含量、净光合速率、植株的干鲜质量, 降低MDA含量和相对电导率。唐秀巧等[24]在小麦灌浆期高温前通过叶面喷施磷酸二氢钾(PDP)和BR均能显著延缓灌浆期叶片衰老, 促进干物质积累。植物在逆境胁迫下会产生活性氧, 破坏细胞膜结构, 阻碍叶绿素合成, 降低SPAD值, 从而破坏光系统, 减少植物干物质积累量[25]。本试验研究结果表明, 在连续冻害胁迫48 h, 外施EBR的处理组小麦MDA含量和相对电导率显著降低, 而Pro含量显著提高, SPAD值、籽粒产量、千粒质量和地上部干物质质量均显著高于对照, 与前人研究结果一致。
在植物逆境研究中, 前人的研究表明, 抗氧化酶SOD、POD、CAT 等活性的高低与植物的抗逆能力密切相关[26], 但植物体内其他的水解酶的存在, 极易影响抗氧化酶的提取与活性的测定, 而进行转录水平的检测则受其他水解酶的影响较小。在前期研究中, 本课题组筛选出与冻害胁迫相关的5个抗逆基因SOD、POD、CAT、P5CS和WCS120。其中, SOD基因控制超氧化物歧化酶的合成, 超氧化物歧化酶作为清除植物体内活性氧(ROS)的清除剂, 是首个响应逆境胁迫并参与反应的酶[12]; POD 基因控制过氧化物酶的合成, 过氧化物酶能分解植物体内的H2O2, 降低其对细胞的危害[13]; CAT基因控制过氧化氢酶的合成, 过氧化氢酶是一种强抗氧化酶, 催化H2O2分解成水和氧, 在植物对非生物胁迫的响应中起着关键作用[14]; WCS120基因是一种受低温特异性诱导的基因, 其编码的蛋白质被认为在小麦的冷驯化过程中起着重要的作用, WCS120蛋白的积累量与高抗冻冬小麦组织的耐寒性呈正相关[16]。吡咯琳-5-羧酸合成酶(Δ 1-pyrroline-5-carboxylate synthetase, P5CS)是脯氨酸合成中谷氨酸合成途径的关键酶[15], Pro含量和植物的逆境耐受力呈正相关[27], 植物体内的Pro含量与P5CS基因的相对表达量呈正相关[28, 29]。因此, SOD、POD、CAT、P5CS和WCS120这5个抗逆相关基因的相对表达量可作为小麦抗冻性的参考依据。本试验研究结果表明, 在冻害胁迫处理48 h, 对照组SOD基因的相对表达量较冻害胁迫处理前无显著变化, POD、P5CS和WCS120均显著升高, 而外施EBR的处理组小麦植株, 除SOD外POD、CAT、P5CS和WCS120的表达量较冻害胁迫处理前均显著提高, 在冻害胁迫48 h 5个抗逆基因的相对表达量均显著高于对照。
在冻害胁迫条件下, 抗逆基因转录水平的检测结果与生理指标鉴定结果一致, 表明在冻害胁迫下, 外施EBR可提高小麦植株的抗氧化酶活性, 增加其清除活性氧的能力, 降低膜脂过氧化程度; 同时P5CS基因的高表达表明, 外施EBR可提高植株体内游离脯氨酸含量, 增加小麦的抗寒性。SPAD值的显著提高表明, 外施EBR可缓解冻害胁迫下小麦叶片衰老, 延长其功能期, 提高小麦植株物质积累量; 幼穗受冻率的检测结果同样表明, 在小麦拔节期外施EBR可降低其幼穗受冻率, 提高籽粒产量和千粒质量。因此, EBR拔节期外施可起到缓解冻害的效应, 从而提高小麦的抗冻性。基于此, 在春季低温冻害来临之际, 可以通过施用EBR减轻低温冻害对冬小麦生产造成的产量损失。
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|